- Óxido nítrico sintasa
-
La óxido nítrico sintasa es la enzima responsable de transformar la L-arginina en óxido nítrico (NO) en una catálisis orgánica de oxidorreducción (reacción RedOx), sin gasto de energía o ATP.
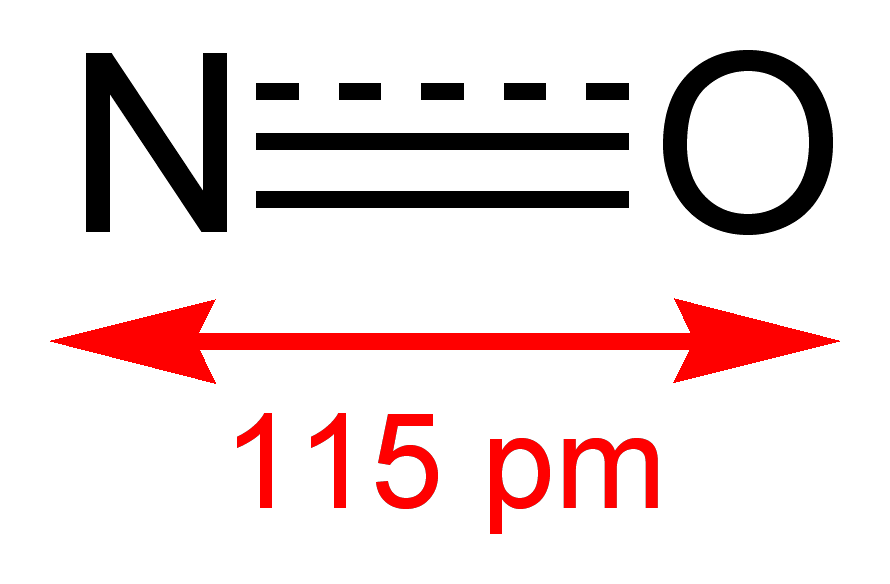
 Óxido nítrico
Óxido nítrico
 El mecanismo de reacción de la "enzima NOS".
El mecanismo de reacción de la "enzima NOS".
Se identifica la enzima en coloreado.Esta sintasa (sigla en inglés NOS; Número EC: 1.14.13.39) es una oxidorreductasa (ya que tiene un dominio oxidasa y un dominio reductasa) responsable de la síntesis de óxido nítrico (ON, siglas en español, y NO, siglas en inglés y más aceptado por poseer el orden de electronegatividad creciente) a partir del átomo terminal de nitrógeno de la L-arginina en presencia de nad-fosfato reducido o NADPH (nicotinamida-adenín-dinucleótido fosfato reducido) y dioxígeno (O2). NOS es la única enzima conocida que une FAD (flavín-adenín-dinucleótido), FMN (flavín mononucleótido), hemo, tetrahidrobiopterina (BH4) y calmodulina.
Contenido
Función
La sintasa produce óxido nítrico (NO) catalizando la oxidación de cinco electrones del nitrógeno guanidina del aminoácido L-arginina (L-Arg).
La oxidación de L-Arg a L-citrulina ocurre vía dos sucesivas reacciones de monooxigenación, produciendo Nω-hidroxi-L-arginina (NOHLA) como intermediario. Dos moles de O2 y 1,5 moles de NADPH se consumen por mol de NO formado.
- L-Arg + NADPH + H+ + O2 → NOHLA + NADP+ + H2O
- NOHLA + ½ NADPH + ½ H+ + O2 → L-citrulina + ½ NADP+ + NO + H2O
Nota sobre la nomenclatura
Es común escribir erróneamente
 óxido nítrico sintetasa, que no existe, en lugar de sintasa, que es lo correcto en este caso. Las sintetasas (en inglés synthetases) son una categoría de enzimas (categoría EC 6) que catalizan la unión entre dos moléculas de sustrato.[1] Son conocidas como ligasas, y su reacción de síntesis requiere la hidrólisis de un nucleótido trifosfato (como ATP).[2]
óxido nítrico sintetasa, que no existe, en lugar de sintasa, que es lo correcto en este caso. Las sintetasas (en inglés synthetases) son una categoría de enzimas (categoría EC 6) que catalizan la unión entre dos moléculas de sustrato.[1] Son conocidas como ligasas, y su reacción de síntesis requiere la hidrólisis de un nucleótido trifosfato (como ATP).[2]Estrictamente las sintetasas (ligasas) son aquellas enzimas que necesitan ATP para cumplir su función, mientras que las sintasas (oxidorreductasas) no requieren de él, como en este caso.
Clasificación
Óxido nítrico sintasa 1 (neuronal) 
HUGO 7872 Símbolo NOS1 Símbolos alt. nNOS Datos genéticos Código de gen NOS1 Tipo de gen Gen Codificante Locus Cr. 12 q14-qter Estructura/Función proteica Tamaño 1434 (aminoácidos) Bases de datos Número EC 1.14.13.39 Entrez 4842 OMIM 163731 RefSeq NM_000620 UniProt P29475 Óxido nítrico sintasa 2 (inducible) HUGO 7873 Símbolo NOS2 Símbolos alt. iNOS Datos genéticos Código de gen NOS2 Tipo de gen Gen Codificante Locus Cr. 17 q11.2-q12 Estructura/Función proteica Tamaño 1153 (aminoácidos) Bases de datos Número EC 1.14.13.39 Entrez 4843 OMIM 163730 RefSeq NM_000625 UniProt P35228 Óxido nítrico sintasa 3 (endotelial) HUGO 7876 Símbolo NOS3 Símbolos alt. eNOS Datos genéticos Código de gen NOS3 Tipo de gen Gen Codificante Locus Cr. 7 q36 Estructura/Función proteica Tamaño 1203 (aminoácidos) Bases de datos Número EC 1.14.13.39 Entrez 4846 RefSeq NM_000603 UniProt P29474 Los distintos miembros de la familia NOS están codificadas por diferentes genes.[3] NOS es una de las enzimas más regulados en la biología. Hay tres isoformas conocidas, dos son constitutivas (CNOS) y la tercera es inducible (iNOS).[4] La clonación de enzimas NOS señala que NOS2 es inducible, mientras que NOS3 y NOS1 son constitutivos, en cerebro y en tejido endotelial respectivamente.[5] Recientemente, la actividad de NOS se ha demostrado en varias especies bacterianas, incluyendo los mismos agentes patógenos conocidos como Bacillus anthracis y Staphylococcus aureus.[6] [7] Se ha demostrado que la NOS bacteriana (bNOS) protege contra el estrés oxidativo, los antibióticos diversos y la respuesta inmune.[8] [9] Las diferentes formas de NO sintasa han sido clasificados de la siguiente manera:
nombre Gen(es) Ubicación Función NOS neuronal (nNOS ó NOS1) NOS1 - comunicación celular
NOS inducible (iNOS ó NOS2) NOS2A - sistema inmune
- sistema cardiovascular
- Defensa inmune contra patogenos
NOS endotelial (eNOS ó NOS3 ó cNOS) NOS3 NOS bacteriana (bNOS) multiple - Varias especies Gram(+)
- Defensa contra estres oxidativo, antibioticos y ataque inmune.
nNOS
NOS neuronal (nNOS) produce NO en tejido nervioso, tanto central como periférico. NOS neuronal también realiza un papel en la comunicación celular y se asocia con la membrana plasmática. La acción de nNOS puede ser inhibida por NPA (N-propil-L-arginina). Esta forma de la enzima se inhibe específicamente por 7-nitroindazol.[10]
iNOS
A diferencia de la regulación crítica dependiente de calcio de las enzimas NOS constitutivas (nNOS y eNOS), iNOS ha sido descrito como insensible a calcio, probablemente debido a su interacción estrecha no covalente con calmodulina (CAM) y Ca2+. Aunque las pruebas de 'linea de base' la expresión de iNOS ha sido difícil de alcanzar, IRF-1 y la activación dependiente de NF-kB del promotor inducible NOS apoya la idea de una estimulación de la transcripción mediada por inflamación.
Desde una perspectiva funcional, es importante reconocer que la inducción de la iNOS de alto rendimiento por lo general ocurre en un ambiente oxidativo, por lo que los altos niveles de NO tienen la oportunidad de reaccionar con [especies reactiva de nitrógeno|superóxido] que conducen a la formación de peroxinitrito y toxicidad celular.
Estas propiedades pueden definir las funciones de la iNOS en la inmunidad del huésped, lo que permite su participación en actividades antimicrobianas y antitumorales como parte de la explosión oxidativa de los macrófagos.[11]
Se ha sugerido que la generación patológica de óxido nítrico mediante el aumento de producción de iNOS puede disminuir el batido ciliar de las trompas y las contracciones del músculo liso y, por tanto afectan el transporte de embriones, que por lo tanto puede resultar en embarazo ectópico.[12]
eNOS
NOS endotelial (eNOS), también conocido como óxido nítrico sintasa 3 (NOS3), genera NO en los vasos sanguíneos y está involucrada con la regulación de la función vascular. Un NOS constitutivo dependiente de Ca2+ proporciona una liberación basal de NO. eNOS se asocia con la membrana plasmática que rodea las células y las membranas del reticulo de Golgi dentro de las células.
Estructura
NOSs puede ser un dimero, calmodulina-dependiente ó hemoproteína tipo citocromo p450 conteniendo calmodulina que combina dominios catalíticos reductasa y oxigenasa en un dímero, portando flavina adenina dinucleótido (FAD) y flavina mononucleótido (FMN), y lleva a cabo una oxidación 5` de una arginina con la ayuda de tetrahidrobiopterina.[13]
Los tres isoformas (cada una de las cuales se presume que funcionan como un homodímero) durante la activación comparten un dominio reductasa carboxilo-terminal homóloga a la citocromo P450 reductasa. También comparten un dominio oxigenasa amino-terminal, que incluye un grupo prostético hemo, que está vinculado en el centro de la proteína a un dominio de unión a calmodulina. La unión de la calmodulina parece actuar como un "interruptor molecular" para permitir el flujo electrónico de grupos prostéticos de flavina del dominio reductasa al hemo. Esto facilita la conversión de O2 y L-arginina a NO y L-citrulina. El dominio oxigenasa de cada isoforma NOS también contiene un grupo prostético BH4, que se requiere para la generación eficiente de NO. A diferencia de otras enzimas donde BH4 se utiliza como fuente de equivalentes de reducción y reciclado por dihidrobiopterina reductasa, BH4 activa el hemo unido al O2 donando un único electron, que luego se recaptura para permitir la liberación de óxido nítrico.
La primer óxido nítrico sintasa identificada fue encontrada en el tejido neuronal (nNOS ó NOS1), la NOS endotelial (eNOS ó NOS3) fue la tercera en ser identificada. Estas fueron clasificados originalmente como "constitutiva expresada" ("constitutively expressed") y "sensible a Ca2+", pero ahora se sabe que están presentes en diferentes tipos de células y que su expresión es regulada en virtud de determinadas condiciones fisiológicas.
Para NOS1 y NOS3, concentraciones fisiológicas de Ca2+ en las células regulan la unión de la calmodulina a los dominios "traba" ó "latch", iniciando así la transferencia de electrones desde el grupo flavin al hemo. Por el contrario, la calmodulina sigue siendo fuertemente unida a la isoformainducible y insensible a Ca2+ (iNOS ó NOS2), incluso en un nivel bajo intracelular de Ca2+, actuando esencialmente como una subunidad de esta isoforma.
El óxido nítrico puede regular por sí mismo la expresión y actividad de NOS. En concreto, se ha demostrado que NO desempeña un importante rol regulador negativo (feedback negativo) sobre NOS3 y por consiguiente en la función de las células endoteliales vasculares. Este proceso, conocido oficialmente como S-nitrosación (s-nitrosation), y mencionado por muchos en el campo como S-nitrosilación (s-nitrosilation), ha demostrado inhibir de forma reversible la actividad NOS3 en las células endoteliales vasculares. Este proceso puede ser importante ya que está regulado por las condiciones redox de la celula y por tanto, puede proporcionar un mecanismo para la asociación entre el "estrés oxidativo" y la disfunción endotelial. Además de NOS3, tanto NOS1 y NOS2 se han encontrado S-nitrosados, pero la evidencia para la regulación dinámica de las isoformas de NOS por este proceso es menos completa. Además, se ha demostrado que tanto NOS1 y NOS2 forman complejos ferrosos nitrosilo en los grupos hemo, que pueden actuar parcialmente para auto-inactivar estas enzimas en determinadas condiciones. El paso limitante para la producción de óxido nítrico también puede ser la disponibilidad de L-arginina en algunos tipos de células. Esto puede ser particularmente importante después de la inducción de NOS2.
Referencias
- ↑ Nomenclature Commission of International Union of Biochemistry and Molecular Biology. Newsletter 1984.
- ↑ Subiza, J. L. (2002). «Carta al Director. Sintetasas y sintasas». Alergología e Inmunología Clínica 17: pp. 188. http://revista.seaic.es/junio2002/188.pdf. Consultado el 20 de octubre de 2008.
- ↑ Tylor BS, Kion YM, Wang Qi, Sharpio RA, Billiar TR y Geller D.A. (1997) "Nitric oxide down regulates hepatocyte-inducible nitric oxide synthase gene expression", Arco. Surg. 1, (32). Noviembre, 1177-1182
- ↑ Majano P. Lara-Pezzi E., López-Cabrera M., Apolinario A., Moreno-Otero R. y García-Monzón C. (2001): Hepatitis B virus X protein transactivates inducible nitric oxide synthase gene promoter through the proximal nuclear factor kappa B-binding site: evidence that cytoplasmic location of X protein is essential for gene transactivation.Hepatology Dec; 34(6): 1218-1224.)
- ↑ Gilkeson G., J. Mudgetts, Seldin MC, Ruiz P., Alexander A., MA Misukonis, DS Pistesky JB y Weinberg (1997): Clinical and serologic manifestation of autoimmune disease in MRL-Mice lacking Nitric Oxide Synthase Type 2. J. Exp. Med. 186 3): Agosto, 4: 365-373.
- ↑ Gusarov I, H Starodubtseva, ZQ Wang, L McQuade, SJ Lippard, DJ Stuehr y Nudler E. Bacterial nitric-oxide synthases operate without a dedicated redox partner. J Biol Chem. 2008 May 9;283(19):13140-7
- ↑ Shatalin K, Gusarov I, E Avetissova, Y Shatalina, LE McQuade, Lippard SJ, y Nudler Bacillus anthracis-derived nitric oxide is essential for pathogen virulence and survival in macrophages. Proc Natl Acad Sci U S A. 2008 Jan 22;105(3):1009-13
- ↑ me Gusarov, E. Nudler. NO-mediated cytoprotection: instant adaptation to oxidative stress in bacteria. Proc Natl Acad Sci U S A. 2005 Sep 27;102(39):13855-60
- ↑ Gusarov I, K Shatalin, M Starodubtseva y Nudler E. Endogenous nitric oxide protects bacteria against a wide spectrum of antibiotics. Science. 2009 Sep 11;325(5946):1380-4
- ↑ Southan GJ, Szabó C (Febrero). Biochem. Pharmacol. 51: pp. 383 a 94. doi:. PMID 8619882.
- ↑ Mungrue, et al., Heart Fail 2002 Rev. octubre 7; (4):407-22.
- ↑ Al-Azemi M, Refaat B, S Amer, Ola B, N Chapman, Ledger W (Mayo 2009). Fértil. Steril. 94: pp. 833 a 40. doi:.
- ↑ Chinje CE y IJ Stratford (1997): Role of nitric oxide in growth of solid tumors: a balancing act. Essays Biochem; 32: 61-72.
Categorías:- Genes del cromosoma 12
- Genes del cromosoma 17
- Genes del cromosoma 7
- Oxidorreductasas
Wikimedia foundation. 2010.