- Flavonoide
-
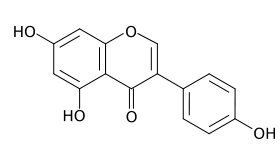
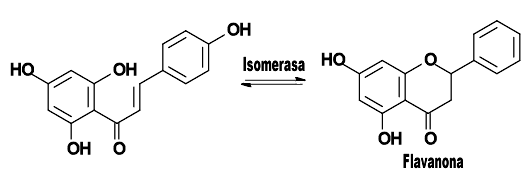
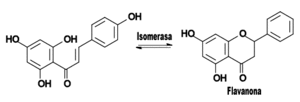 La estructura base de los flavonoides tiene el esqueleto de una chalcona, y la acción de la enzima isomerasa la convierte en una flavanona.
La estructura base de los flavonoides tiene el esqueleto de una chalcona, y la acción de la enzima isomerasa la convierte en una flavanona.
 Esqueleto de los isoflavonoides..
Esqueleto de los isoflavonoides..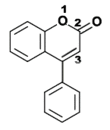 Esqueleto de los neoflavonoides.
Esqueleto de los neoflavonoides.Flavonoide (del latín flavus, "amarillo") es el término genérico con que se identifica a una serie de metabolitos secundarios de las plantas. Son sintetizados a partir de una molécula de fenilalanina y 3 de malonil-CoA, a través de lo que se conoce como "vía biosintética de los flavonoides", cuyo producto, la estructura base, se cicla gracias a una enzima isomerasa. La estructura base, un esqueleto C6-C3-C6, puede sufrir posteriormente muchas modificaciones y adiciones de grupos funcionales, por lo que los flavonoides son una familia muy diversa de compuestos, aunque todos los productos finales se caracterizan por ser polifenólicos y solubles en agua. Los flavonoides que conservan su esqueleto pueden clasificarse, según las isomerizaciones y los grupos funcionales que les son adicionados, en 6 clases principales: las chalconas, las flavonas, los flavonoles, los flavandioles, las antocianinas, y los taninos condensados.[1] más una séptima clase, las auronas, tenidas en cuenta por algunos autores por estar presentes en una cantidad considerable de plantas. También el esqueleto puede sufrir modificaciones, convirtiéndose entonces en el esqueleto de los isoflavonoides o el de los neoflavonoides, que por lo tanto también son derivados de los flavonoides.
Los flavonoides se biosintetizan en todas las "plantas terrestres" o embriofitas, y también en algunas algas Charophyta, y aunque todas las especies comparten la vía biosintética central, poseen una gran variabilidad en la composición química de sus productos finales y en los mecanismos de regulación de su biosíntesis, por lo que la composición y concentración de flavonoides es muy variable entre especies y en respuesta al ambiente. Los flavonoides son sintetizados en el citoplasma y luego migran hacia su destino final en las vacuolas celulares. Cumplen funciones metabólicas importantes en las plantas, algunas funciones son comunes a todas las plantas y otras son específicas de algunos taxones. Como ejemplo de funciones universales, los flavonoides son responsables de la resistencia de las plantas a la fotooxidación de la luz ultravioleta del Sol, intervienen en el transporte de la hormona auxina, y se cree que funcionan como defensa ante el herbivorismo. Una función importante cumplida en muchas plantas es la atracción de los animales polinizadores, a través del color o el olor que dan a la planta o a sus flores.
Los flavonoides han adquirido notoriedad pública a raíz de su actividad biológica en el hombre, que los consume con los vegetales. Los flavonoides poseen propiedades muy apreciadas en medicina, como antimicrobianos, anticancerígenos, disminución del riesgo de enfermedades cardíacas, entre otros efectos. También son conocidos por los cultivadores de plantas ornamentales, que manipulan el ambiente de las plantas para aumentar la concentración de flavonoides que dan el color a las hojas y a las flores.
Debido a las importantes funciones metabólicas que los flavonoides tienen en las plantas y los animales, sus vías biosintéticas y mecanismos de regulación están siendo cuidadosamente estudiados. La ciencia aplicada aprovechó este conocimiento en muchos trabajos de ingeniería metabólica, en los que se buscó por ejemplo, aumentar la concentración de flavonoides beneficiosos en las plantas de consumo humano o de uso farmacéutico, modificar su concentración en flores ornamentales para cambiarles el color, e inhibir su producción en el polen para lograr la esterilidad de los híbridos de interés comercial. En lo que respecta a su producción, se ha desarrollado con éxito un cultivo de bacterias que sintetiza flavonoides de interés humano.
Los científicos dieron usos variados a los flavonoides: los genes de la biosíntesis de flavonoides fueron usados como herramienta para analizar los cambios en el ADN, son ejemplos conocidos el descubrimiento de las leyes de Mendel (que pudo rastrear la herencia de los genes de los flavonoides que dan el color a los guisantes), y el descubrimiento de los genes saltarines de Barbara McClintock (que al "saltar" hacia un gen de un flavonoide lo inutilizan y no se expresa el color en el grano de maíz). La extracción e identificación de flavonoides también fue muy usada por los botánicos sistemáticos para establecer parentescos entre especies de plantas.
Aún queda mucho por investigar de los flavonoides, de su valor medicinal, y de su impacto en la nutrición y la salud humana y de los animales. También es necesario continuar la investigación de su estructura, su metabolismo y su biodisponibilidad, por lo que se esperan importantes progresos en este campo[2]
Contenido
Descubrimiento
Probablemente la primera vez que la ciencia describió a los flavonoides fue cuando Robert Boyle en 1664 hizo una primera descripción de los efectos de los pigmentos de las flores en medio ácido y en medio básico.[1]
El primer flavonoide fue identificado en 1930 por el premio Nobel de Fisiología y Medicina Szent-Györgyi, quien aisló de la cáscara de limón una sustancia, la citrina, que probó regular la permeabilidad de los capilares al ser consumida.
Los flavonoides se denominaron al principio vitamina P (por permeabilidad) y también vitamina C2, porque algunos tenían propiedades similares a la vitamina C. El hecho de que los flavonoides fueran vitaminas no pudo ser confirmado, y ambas denominaciones se abandonaron alrededor de 1950.[3]
Estructura química
Flavonoide base y la acción de la enzima isomerasa.El primer flavonoide sintetizado por la "vía biosintética de los flavonoides" es una chalcona, cuyo esqueleto es un anillo bencénico unido a una cadena propánica que está unida a su vez a otro anillo bencénico. En la mayoría de los flavonoides, la cadena de reacciones continúa, por lo que la cadena carbonada que une los anillos aromáticos se cicla por acción de una enzima isomerasa, creando una flavanona.
Muchas veces la biosíntesis continúa y los productos finales, también flavonoides, quedan unidos a muy diversos grupos químicos, por ejemplo los flavonoides glucosidados portan moléculas de azúcares o sus derivados. También pueden encontarse flavonoides parcialmente polimerizados dando lugar a dímeros, trímeros, o complejos multienlazados, como los taninos condensados.Todos los flavonoides poseen las características de ser polifenólicos y solubles en agua. Poseen un máximo de absorción de luz a los 280 nm.
Extracción y análisis
La extracción de los flavonoides se realiza con solventes orgánicos de alta polaridad como el etanol.(el acetato de etilo se usa con bastante precaución ya que por calentamiento puede reaccionar con ciertos compuestos). Posteriormente se realizan extracciones sucesivas con solventes de polaridad creciente como: Hexano / Cloroformo (para flavonoides de baja polaridad). Acetato de etilo (para flavonoides de mediana polaridad). Butanol (para flavonoides de alta polaridad y por lo general los de mayor uso farmacológico gracias a la presencia de grupos funcionales de tipo hidroxilo y carbonilo)
La espectrofotometría es útil para analizar la concentración de flavonoides en una sustancia.
Muchas veces esa medida se realiza acoplada a una separación cromatográfica como por ejemplo HPLC.
Clasificación de los flavonoides
De acuerdo con la nomenclatura de la Unión Internacional de Química Pura y Aplicada.[4] pueden clasificarse, según su esqueleto y vía metabólica, en:
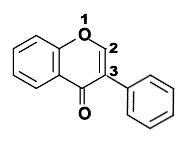
- Flavonoides, derivados de la estructura 2-fenilcromen-4-ona (2-fenil-1,4-benzopirona).
- Isoflavonoides, derivados de la estructura 3-fenilcromen-4-ona (3-fenil-1,4-benzopirona).
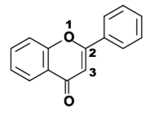
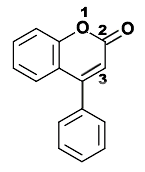
- Neoflavonoides, derivados de la estructura 4-fenilcumarina (4-fenil-1,2-benzopirona).
 Estructura química de la 2-fenilcromen-4-ona (2-fenil-1,4-benzopirona), esqueleto de los flavonoides.
Estructura química de la 2-fenilcromen-4-ona (2-fenil-1,4-benzopirona), esqueleto de los flavonoides.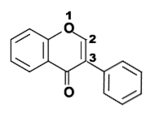 Estructura química de la 3-fenilcromen-4-ona (3-fenil-1,4-benzopirona), esqueleto de los isoflavonoides..
Estructura química de la 3-fenilcromen-4-ona (3-fenil-1,4-benzopirona), esqueleto de los isoflavonoides.. Estructura química de la 4-fenilcumarina (4-fenil-1,2-benzopirona), esqueleto de los Neoflavonoides.
Estructura química de la 4-fenilcumarina (4-fenil-1,2-benzopirona), esqueleto de los Neoflavonoides.Los isoflavonoides se forman por migración de un anillo bencénico de la posición 2 a 3 del anillo central. El grupo integra más de 230 estructuras, y los dos más conocidos son la genisteína y la daidzeina[5] Su función es defender a las plantas del ataque de patógenos.
Dentro de los flavonoides, se reconocen 6 y quizás 7 clases principales, según los grupos funcionales que posean: las chalconas, las flavonas, los flavonoles, los flavandioles, las antocianinas, los taninos condensados, y algunos autores consideran también a las auronas, que otros integran a las chalconas. También hay otros derivados de los flavonoides que poseen modificaciones tales que no entran dentro de ninguna de estas clases principales.
El número de flavonoides diferentes que es en teoría posible es astronómico, si se tiene en cuenta que diez de los carbonos del esqueleto del flavonoide pueden ser sustituidos por una variedad de grupos diferentes, que a su vez pueden ser hidroxilados, metoxilados, metilados, isoprenilados o benzilados. Además, cada grupo hidroxilo y algunos de los carbonos pueden ser sustituidos por uno o más azúcares diferentes, y a su vez, cada uno de esos azúcares puede ser acilado con una variedad de ácidos fenólicos o alipáticos diferentes. Se han identificado y aislado alrededor de 9.000 flavonoides, pero sin duda aún hay muchos más por descubrir.[6]
1. Las chalconas están implicadas en la estimulación de la polinización gracias a que inducen el desarrollo de colores en el espectro de lo visible y en el UV que atraen a insectos (mariposas y abejas)
 Estructura molecular de la flavona
Estructura molecular de la flavona2. Las flavonas son amarillas y pueden estar en algunas flores, como en la prímula, dándoles un color amarillo a sus pétalos, o en frutos, como en la piel de las uvas, son las responsables del color amarillento de los vinos blancos. Hay tres flavonas importantes: la tricetina, presente en el polen de algunas mirtáceas, y también en las podocarpáceas (Podocarpus spp.); apigenina, presente en muchas plantas como la camomila, (Matricaria recutita) o el espino blanco (Crataegus laevigata), da un color marrón marfileño a las flores si se presenta sola; y luteolina, de color amarillo, que incluso sirve para teñir lana y otros tejidos, para lo cual se ha empleado la Retama de los tintoreros (Genista tinctoria).
3. Los flavonoles suelen ser incoloros o amarillos y se encuentran en las hojas y en muchas flores. Los más importantes son tres: quercetina, es el flavonol amarillo del polen de muchas fagáceas (Quercus sp.);[7] miricetina, presente en la uva; y kaempferol, está presente en las inflorescencias y las protege de la luz ultravioleta. La fisetina es un flavonol que se extrae de la planta del género Amphipterygium.
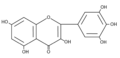 Estructura molecular de la miricetina.
Estructura molecular de la miricetina.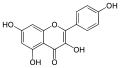 Estructura molecular del kaempferol.
Estructura molecular del kaempferol.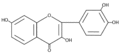 Estructura molecular de la fisetina.
Estructura molecular de la fisetina.4. Hay tres flavandioles característicos: leucocianidina, presente en algunas plantas, como en el plátano, o en el muérdago criollo (Ligaria cuneifolia); leucopelargonidina, presente como tal en cierta concentración en la alfalfa de secano (Medicago truncatula); y leucodelfinidina, que es activa en el castaño de indias (Aesculus hippocastanum).
5. Las antocianinas, son los pigmentos hidrosolubles presentes en el líquido vacuolar de las células responsables de la mayoría de las coloraciones rojas, azules y violetas de las flores y hojas.[8]
6. Los taninos condensados son macromoléculas constituidas por unidades de flavonoides llamadas antocianidina. Los taninos están muy ampliamente distribuidos en las plantas como en el té, donde contribuyen al sabor astringente.[9]
7. Las auronas son responsables de la coloración de algunas plantas. A pesar de que se ha sugerido que estos compuestos están relacionados estrechamente con las chalconas, hay pocos indicios acerca de sus vías biosintéticas[10]
8. Las flavanonas son precursores de otros flavonoides más complejos, pero se encuentran como tales en altas concentraciones en los cítricos. Las más importantes son naringenina, presente en el zumo de naranja, limón o pomelo, dándole un sabor amargo; liquiritigenina, presente en el regaliz; y eriodictiol, se presenta en el guisante actuando como quimioatrayente para interactuar con agrobacterias.
9. Los dihidroflavonoles son los precursores directos de flavandioles y flavonoles, pero también tienen cierta actividad como tales en algunas plantas. Hay tres importantes: dihidromiricetina, presente en las partes aéreas de los brezos (Erica spp.), dihidroquercetina, en las uvas blancas o en la zarzaparrilla (Smilax aristolochiaefolia); y dihidrokaempferol.
Los flavonoides en las plantas
Origen evolutivo
Los flavonoides aparecieron por primera vez en los ancestros de las embriofitas, que comprende al grupo monofilético de todas las plantas terrestres (musgos, helechos, gimnospermas y angiospermas). Se cree que fueron una de las adaptaciones clave para la transición a la vida terrestre desde el alga verde ancestral, debido a su capacidad de absorber la radiación ultravioleta, mucho más intensa en la atmósfera que en el agua.[11]
Las enzimas de la biosíntesis de los flavonoides aparentemente derivaron de enzimas del metabolismo primario de las plantas, que tenían genes duplicados, lo que habrá permitido la adaptación de algunas esas enzimas a otras funciones específicas (Iida et al. 1999[12] Rausher et al. 1999,[13] Durbin et al. 2000.[14] ).
La vía biosintética de los flavonoides se ha conservado enormemente en el transcurso de la evolución de las plantas, pero ha habido considerable divergencia tanto en los roles que fueron cumpliendo sus productos finales, como en los mecanismos que regulan su expresión.[15]
Biosíntesis
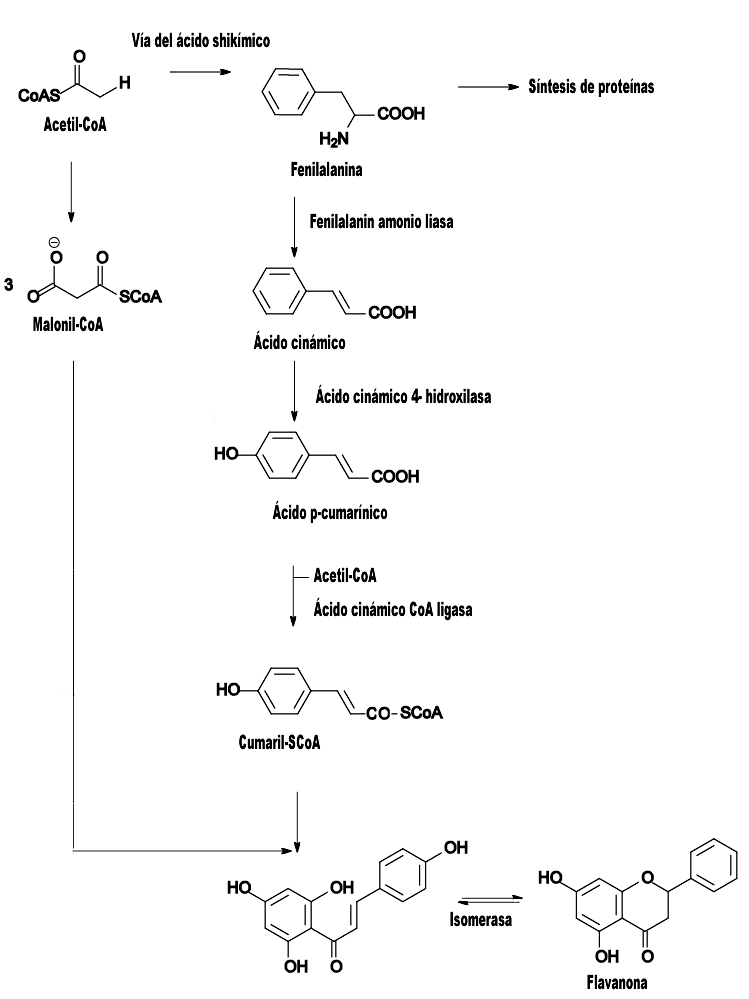
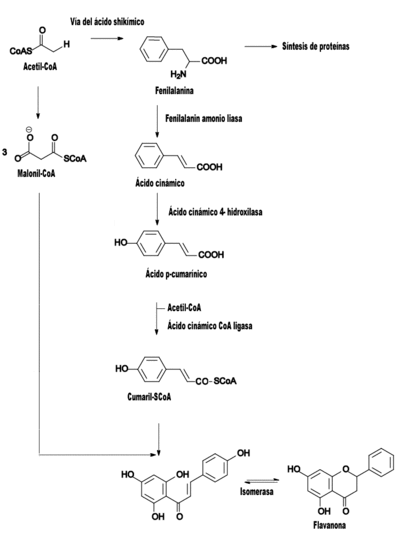 Ruta de biosíntesis de los flavonoides en las plantas.
Ruta de biosíntesis de los flavonoides en las plantas.La vía del ácido shikímico se inicia en los plastos por condensación de dos productos fotosintéticos, la eritrosa 4-P con el fosfoenolpiruvato (PEP), y por diversas modificaciones se obtiene el ácido shikímico, del cual derivan directamente algunos fenoles en los vegetales. Pero la vía del ácido shikímico normalmente prosigue, y la incorporación de una segunda molécula de PEP conduce a la formación de fenilalanina.
La vía biosintética de los flavonoides comienza cuando la fenilalanina, por acción de la enzima fenilalanina amonioliasa (PAL) se transforma en ácido cinámico, que luego es transformado en ácido p-cumarínico por incorporación de un grupo hidroxilo a nivel de anillo aromático, y la acción de una CoA ligasa lo transforma en cumaril-SCoA, el precursor de la mayoría de los fenoles de origen vegetal, entre los que se encuentran los flavonoides[9]
Regulación de la biosíntesis
- La vía del ácido shikímico es dependiente de la luz.
- La acción de la fenilalanina amonioliasa, que inicia la vía biosintética de los flavonoides, es fundamental para la vida de las plantas y por ello está estrictamente regulada. Entre otros factores, la fenilalanina amonioliasa es activada por la luz, y depende además de la concentración de diferentes hormonas vegetales. La actividad de la fenilalanina amonioliasa suele aumentar cuando a los vegetales se les somete a situaciones de estrés, como puede ser la falta de agua ("estrés hídrico"), infecciones fúngicas o bacterianas, radiaciones UV, y el frío (por esto último las plantas sometidas a bajas temperaturas suelen presentar coloraciones rojizas en tallos y hojas, y cuando los inviernos son muy fríos, las flores desarrollan colores muy intensos en la primavera siguiente).
- Hay isozimas dedicadas a la producción de flavonoides diferentes en respuesta a señales ambientales diferentes.[16]
Funciones en las plantas
- Protección ante la luz UV. Los flavonoides incoloros suelen acumularse en las capas más superficiales de las plantas y captan hasta el 90% de las radiaciones UV, impidiendo los efectos nocivos de estas radiaciones en los tejidos internos.
- Defensa ante el herbivorismo. Algunos flavonoides como los taninos, protegen a las plantas generando sabores desagradables para los herbívoros, principalmente amargos, o texturas que pueden resultar desagradables para los herbívoros, que se ven estimulados a elegir otras plantas.[9]
- Regulación del transporte de la hormona auxina. Las plantas mutantes que no poseen la enzima chalcona sintasa, que forma parte de la vía biosintética de los flavonoides, muestran un crecimiento irregular debido a una deficiencia en el transporte de auxina a través de la planta. Probablemente esa deficiencia se deba a la ausencia de ese flavonoide en la planta mutante[17]
 Flor de Tillandsia aeranthos. El pigmento que le da el color a sus brácteas es un flavonoide.
Flor de Tillandsia aeranthos. El pigmento que le da el color a sus brácteas es un flavonoide.- Atracción de animales polinizadores. Muchos flavonoides son componentes de pigmentos de las flores y hojas que confieren coloraciones atrayentes de insectos, con lo que la función de muchos flavonoides sería la de atraer a los polinizadores hacia las flores Un caso muy destacado, es el de las bromeliáceas entre las que se encuentran las especies Tillandsia y Billbergia, que desarrollan sus flores sobre un tallo que se elonga sobre una base con hojas en roseta. El tallo elongado está formado por una serie de brácteas que presentan un color rojizo muy fuerte antes de la polinización o durante ésta, y luego se hacen más verdosas.[18]
 Hoja de Dionaea muscipula, una planta carnívora. El color de sus hojas se debe a una antocianina.
Hoja de Dionaea muscipula, una planta carnívora. El color de sus hojas se debe a una antocianina.- Atracción de presas. Las plantas carnívoras, como la Drosera y la Dionaea, poseen antocianinas en sus flores y hojas, que cumplen una función de atracción de los insectos que les sirven de alimento.
- Atracción de animales dispersores de semillas y frutos. Algunos flavonoides confieren aromas y colores a los frutos que los hacen más apetecibles para los herbívoros que se alimentan de ellos, cumpliendo así una función de dispersión de las semillas.
 Estructura molecular de la genisteína.
Estructura molecular de la genisteína.- Inducción de la nodulación por parte de las bacterias fijadoras de nitrógeno. Se ha observado que el eriodictiol y la apigenina-7-O-glucósido exudados por la raíz del guisante (Pisum sativum) inducen la nodulación de la agrobacteria Rhizobium leguminosarum. Se ha visto también que dos isoflavonoides encontradas en exudados de soja, la daidzeina y la genisteína son inductores de los genes de la nodulación de varias cepas de Bradyrhizobium japonicum.
- Protección contra los hongos. La pisatina es un isoflavonoide producido por el guisante en respuesta a la infección por hongos e induce la expresión del gen PDA1 en éstos. La pisatina y el eriodictiol inducen la germinación de las esporas de ciertos hongos.
Aplicaciones de los flavonoides
Aplicaciones en medicina
- Para un review en inglés ver Graf et al. 2005.[19]
Los flavonoides consumidos por el hombre le protegen del daño de los oxidantes, como los rayos UV (cuya cantidad aumenta en verano); la polución ambiental (minerales tóxicos como el plomo y el mercurio); algunas sustancias químicas presentes en los alimentos (colorantes, conservantes, etc). Como el organismo humano no tiene la capacidad de sintetizar estas sustancias químicas, las obtiene enteramente de los alimentos que ingiere.
No son considerados vitaminas.
Al limitar la acción de los radicales libres (que son oxidantes), los flavonoides reducen el riesgo de cáncer, mejoran los síntomas alérgicos y de artritis, aumentan la actividad de la vitamina C, , bloquean la progresión de las cataratas y la degeneración macular, evitan las tufaradas de calor en la menopausia y combaten otros síntomas.
En general el sabor es amargo, llegando incluso a provocar sensaciones de astringencia si la concentración de taninos condensados es muy alta. El sabor puede variar dependiendo de las sustituciones presentadas en el esqueleto llegando incluso a usarse como edulcorantes cientos de veces más dulces que la glucosa.
Sus efectos en los humanos pueden clasificarse en:
- Propiedades anticancerosas: muchos han demostrado ser tremendamente eficaces en el tratamiento del cáncer. Se sabe que muchos inhiben el crecimiento de las células cancerosas. Se ha probado contra el cáncer de hígado[20]
- Propiedades cardiotónicas: tienen un efecto tónico sobre el corazón, potenciando el músculo cardíaco y mejorando la circulación. Atribuidas fundamentalmente al flavonoide quercetina aunque aparece en menor intensidad en otros como la genisteína y la luteolina. Se ha estudiado que los flavonoides reducen el riesgo de enfermedades cardíacas.[21] [22]
- Fragilidad capilar: mejoran la resistencia de los capilares y favorecen el que éstos no se rompan, por lo que resultan adecuados para prevenir el sangrado. Los flavonoides con mejores resultados en este campo son la hesperidina, la rutina y la quercetina.
- Propiedades antitrombóticas: la capacidad de estos componentes para impedir la formación de trombos en los vasos sanguíneos posibilita una mejor circulación y una prevención de muchas enfermedades cardiovasculares.
- Disminución del colesterol: poseen la capacidad de disminuir la concentración de colesterol y de triglicéridos.
- Protección del hígado: algunos flavonoides han demostrado disminuir la probabilidad de enfermedades en el hígado. Fue probado en laboratorio que la silimarina protege y regenera el hígado durante la hepatitis. Junto con la apigenina y la quercetina, son muy útiles para eliminar ciertas dolencias digestivas relacionadas con el hígado, como la sensación de plenitud o los vómitos.
- Protección del estómago: ciertos flavonoides, como la quercetina, la rutina y el kaempferol, tienen propiedades antiulcéricas al proteger la mucosa gástrica.
- Antiinflamatorios y analgésicos: la hesperidina por sus propiedades antiinflamatorias y analgésicas, se ha utilizado para el tratamiento de ciertas enfermedades como la artritis. Los taninos tienen propiedades astringentes, vasoconstrictoras y antiinflamatorias, pudiéndose utilizar en el tratamiento de las hemorroides.[9]
- Antimicrobianos: isoflavonoides, furanocumarinas y estilbenos han demostrado tener propiedades antibacterianas, antivirales y antifúngicas.
- Propiedades antioxidantes: En las plantas los flavonoides actúan como antioxidantes, especialmente las catequinas del té verde. Durante años se estudió su efecto en el hombre, y recientemente (5 de marzo del 2007[23] ) se ha concluido que tienen un efecto mínimo o nulo en el organismo humano como antioxidantes.
Por esas causas son prescriptas las dietas ricas en flavonoides, se encuentran en todas las verduras pero las concentraciones más importantes se pueden encontrar en el brócoli,[22] la soja, el té verde y negro, el vino, y también se pueden ingerir en algunos suplementos nutricionales, junto con ciertas vitaminas y minerales. En los frutos, las mayores concentraciones se encuentran en la piel, por lo que es mejor comerlos sin pelar, debidamente lavados previamente.[18] También es importante destacar que muchos de estos compuestos se encuentran en proporciones variables en los diferentes tipos de vinos, siendo responsables del efecto preventivo que tiene el consumo moderado de vino sobre las enfermedades cardiovasculares, cáncer y otras enfermedades degenerativas.[9] Las mayores concentraciones en el tomate están presentes en el de tipo "cherry", y en la lechuga, en la del tipo "Lollo Rosso".[24] La concentración de los flavonoides también varía mucho entre plantas de la misma especie, por lo que se recomienda el consumo de verduras de buena calidad, y como los flavonoides se estropean con facilidad, es recomendado consumirlas en lo posible crudas, y si se cocinan no se recomienda el uso del microondas ni congelarlas antes de hervirlas,[24]
Aplicaciones en las plantas ornamentales y los frutos comerciales
Las plantas de mayor valor estético tienen mayor demanda en el mercado, por lo que se han aplicado los conocimientos en flavonoides para realzar las coloraciones de las plantas.
La fenilalanina amonioliasa es activada por situaciones de estrés, como puede ser el frío. A bajas temperaturas muchas plantas, como las orquídeas, pueden presentar coloraciones rojizas (o violetas) en hojas que inicialmente eran verdes. Esto es no es más que un mecanismo de defensa, pero puede tener aplicaciones comerciales: si se pulveriza con agua un fruto en las horas de mayor temperatura, se favorece la coloración en las zonas en que no se desarrolla bien el color. Este tratamiento produce, al evaporase el agua, un descenso de temperatura del fruto de hasta 10º C, lo que frenaría temporalmente las reacciones de respiración y favorecería la reacción de síntesis de cianidina produciendo unos frutos de colores más vivos, de más valor comercial.
Investigación en flavonoides
La investigación en flavonoides se puede dividir en dos grandes ramas: la investigación básica, que se ocupa de dilucidar las vías biosintéticas de los flavonoides y sus mecanismos de regulación, y la investigación aplicada, representada por la ingeniería genética de flavonoides, que manipula el ADN de las plantas con fines comerciales. Las dos ramas se interrelacionan de forma estrecha, de forma que por un lado, es necesario conocer las vías biosintéticas y de regulación para que la manipulación del ADN tenga mejores probabilidades de éxito, y por otro lado, la misma manipulación del ADN arroja a veces resultados que ayudan a comprender las vías biosintéticas y de regulación. Las dos ramas se diferencian principalmente en los objetivos propuestos, pero se solapan en algunos de los métodos utilizados.
Un tercer aspecto de la investigación es el uso de las vías biosintéticas y de regulación de los flavonoides como herramienta para experimentación en otras áreas, y el uso de los flavonoides en sí mismos, como herramienta para diferenciar las especies en Botánica Sistemática.
Investigación de las vías biosintéticas y mecanismos de regulación
Las vías biosintéticas básicas fueron dilucidadas a través de la cuidadosa identificación y caracterización de numerosas enzimas que intervienen en la biosíntesis. En esta etapa de la investigación, se aprovechó la utilidad de muchos tejidos de plantas que poseían la característica de poseer enzimas de la síntesis de flavonoides en grandes cantidades y que podían ser aisladas con facilidad. Ejemplos de trabajos de este tipo son las células irradiadas de perejil rizado (Petroselinum hortense) de las que se aisló la chalcona sintasa (Kreuzaler et al. 1979[25] ); los cultivos en suspensión de células de semillas de soja (Glycine max) y poroto (Phaseolus vulgaris) de los que se aisló la chalcona isomerasa (Moustafa y Wong 1967[26] Dixon et al. 1982,[27] ); y las flores de Matthiola incana, petunia (Petunia hybrida), y Dianthus caryophyllus, de las que se aisló la flavanona 3-hidroxilasa, la flavonol sintasa, la flavonoide 3'-hidroxilasa, y la dihidroflavonol reductasa (Forkmann et al. 1980[28] Spribille y Forkmann 1984,[29] Britsch y Grisebach 1986,[30] Stich et al. 1992,[31] ). Estos experimentos dejaron al descubierto que las reacciones químicas que mediaban en la biosíntesis de flavonoides en plantas eran una red compleja, y propulsaron los esfuerzos para aislar los correspondientes genes que codificaban para las enzimas descubiertas.[2]
Históricamente la clonación de genes de la vía de los flavonoides muestra que a mayoría de los genes que codifican para enzimas de la vía principal fueron primero aislados por vías bioquímicas, por ejemplo, por información extraída directamente de las características de la enzima, o el uso de anticuerpos usando como antígeno a la enzima purificada. También fueron útiles los mutantes que resultaban de la inserción de elementos transponibles en los genes implicados en la vía biosintética (procedimiento llamado "etiquetado por transposones" utilizado por primera vez en la historia de la ciencia por Federoff et al. en 1984[32] con el que descubrió el gen que codificaba para una enzima de la vía de los flavonoides). La clonación de genes de la vía de la biosíntesis se hizo mayormente en maíz y petunia, con pocos aportes de otras plantas. La vía de los isoflavonoides, que está presente en las legumbres, fue estudiada recientemente en soja y alfalfa (Medicago sativa), si bien los resultados fueron principalmente basados en aproximaciones bioquímicas. Arabidopsis apareció un poco tarde en la escena de identificación de genes, y fueron útiles principalmente los mutantes, ya que se encontraron mutantes de Arabidopsis de la mayor parte de los genes implicados en la vía biosintética de los flavonoides. El uso de Arabidopsis está llenando vacíos de conocimiento, por ejemplo recientemente se identificó un gen que puede estar implicado en la síntesis de taninos condensados (Devic et al. 1999.[33] ). Al insertar genes de maíz en los mutantes de Arabidopsis se ha comprobado que las enzimas de la vía biosintética de los flavonoides se han conservado en su función en distancias evolutivas enormes (Dong et al. 2001[34] ). Los etiquetados por transposón y por T-DNA en maíz, petunia y Arabidopsis también proveen información largamente esperada sobre los genes envueltos en el transporte de flavonoides del sitio de síntesis en el citoplasma hasta la vacuola (Marrs et al. 1995, Alfenito et al. 1998, Debeaujon et al. 2001).
La identificación de genes que codifican para factores reguladores es más reciente, y se basó casi exclusivamente en el etiquetado de transposones primero, y el etiquetado de T-DNA después. Esto es debido principalmente a que las proteínas reguladoras no se acumulan en cantidades importantes en ningún tejido, como sí lo hacen las de biosíntesis, por lo tanto no son fáciles de realizar las aproximaciones de tipo bioquímico con ellas. También hubo problemas al hacer homologías entre especies, porque se han encontrado secuencias altamente conservadas entre factores de transcripción (por ejemplo los dominios bHLH y myb). Pero una vez se desarrolló el etiquetado por transposones, rápidamente se aislaron los factores reguladores de biosíntesis de flavonoides en maíz, petunia y boca de dragón (Antirrhinum majus). Factores reguladores adicionales fueron aislados posteriormente en Arabidopsis por clonado posicional ("positional cloning") y etiquetado de T-DNA. Una aproximación diferente fue realizada aislando los factores de transcripción del perejil a través de South-western y screening de doble híbrido ("two-hybrid screening", Weisshaar et al. 1991, Rügner et al. 2001). El uso de plantas transgénicas para identificar y caracterizar los factores de regulación han encontrado algunas similitudes, pero también importantes diferencias, en los mecanismos por los que la vía de los flavonoides es regulada en diferentes especies de plantas (Lloyd et al. 1992, Quattrocchio et al. 1998, Uimari y Strommer 1998, Bradley et al. 1999).
La caracterización de la vía de los isoflavonoides en alfalfa y soja ha provisto herramientas para la ingeniería metabólica de la síntesis de isoflavonoides en otras especies de leguminosas[35]
El estudio de las 3-desoxiantocianinas ha echado algo de luz sobre sus vías biosintéticas (ver enlace).
También se ha descubierto un importante rol de los flavonoides en la fertilidad masculina (fertilidad del polen) a través del análisis de mutantes de maíz y de petunia que tenían una mutación en la primera enzima de la biosíntesis de flavonoides. Sin embargo un mutante de Arabidopsis en la misma enzima era totalmente fértil, lo que demuestra que los flavonoides no son requeridos por todas las plantas para la formación del tubo polínico (para review, ver Shirley 1996,[36] ).
También los flavonoides han ayudado a definir la especificidad de huésped de microbios como Rhizobium spp. y Agrobacterium spp. (Rolfe 1988[37] Zerback et al. 1989,[38] ).
Los flavonoides también contribuyen al reconocimiento de la planta huésped por parte de las plantas parásitas como Triphysaria versicolor y Cuscuta subinclusa, pero aparentemente no son requeridos para el parasitismo exitoso de Arabidopsis por parte de Orobanche aegyptiaca (Kelly 1990[39] Albrecht et al. 1999,[40] Westwood 2000.[41] ).
Pero así como los flavonoides cumplen con funciones específicas en especies diferentes, también poseen una serie de roles que se conservan ampliamente, para ello son útiles los modelos como Arabidopsis, que provee información genética y molecular que no está disponible en otras plantas. Arabidopsis además tiene la ventaja de hacer más simple la caracterización de la vía de los flavonoides, porque posee un solo gen para la mayoría de las enzimas que intervienen en la vía, a diferencia de lo que pasa con muchas otras plantas que tienen muchas copias de esos genes. Por lo tanto una mutación en un gen de la vía interrumpe todo el flujo de la biosíntesis, en todos los tejidos y en todas las condiciones ambientales. Un ejemplo de su utilidad fue el uso de algunos mutantes de Arabidopsis para demostrar el rol inequívoco de los flavonoides en proteger a la planta de la radiación UV (Li et al. 1993[42] ). Estos mutantes también proveyeron información sobre la contribución de los flavonoides presentes en la cubierta de la semilla para mantener la dormición de la misma (Debeaujon et al. 2000[43] ). Es más, la largamente controvertida hipótesis de que los flavonoides funcionaban como transporte de la hormona auxina (Jacobs y Rubery 1988[44] ) recibió apoyo de estudios en Arabidopsis (Brown et al. 2001[45] ). En cada caso Arabidopsis ayudó a dar las herramientas para investigar estos mecanismos en otras especies de plantas.
Ingeniería genética de flavonoides
- Véase también: En busca de la rosa azul
Debido a las importantes funciones metabólicas que los flavonoides tienen en las plantas, sus vías biosintéticas están estrictamente reguladas. La ingeniería genética aprovechó esta característica de los flavonoides para hacerlos blanco de muchos trabajos de ingeniería metabólica. La ingeniería metabólica de flavonoides se puede definir como la tecnología que manipula el ADN que interviene en la biosíntesis de flavonoides. El ADN que interviene en la biosíntesis de flavonoides puede dividirse en el que codifica para compuestos estructurales y en el que codifica para proteínas que regulan la transcripción (recientemente se ha descripto que la biosíntesis puede ser regulada al nivel de la transcripción[46] ).
La ingeniería metabólica de flavonoides empezó en 1987[47] y ha sido un área de investigación muy fructífera en la década del '90[48] Muchos de los procedimientos están bajo patente.
La ingeniería genética es cara y costosa, por lo que deben realizarse previamente experimentos destinados a conocer en profundidad las vías metabólicas de la planta a tratar, con el objetivo de maximizar las probabilidades de éxito del tratamiento. Por ejemplo se pueden deducir algunos puntos de sus vías biosintéticas analizando la reacción de la planta ante la presencia de ciertos flavonoides conocidos.
Algunos ejemplos de aplicación exitosa de la ingeniería metabólica a los flavonoides son (Forkmann y Martens 2001.[49] ):
- Coloración de las flores. La innovación en el color de las flores de las plantas ornamentales, en especial la oferta de variedades azules o amarillas inexistentes en la naturaleza, es uno de los mayores atractivos que pueden ofrecer los cultivares. Como ejemplo se puede mencionar la Petunia de flores naranjas (el color es concedido por flavonoides sintetizados gracias a genes traídos de otras plantas), y los claveles violetas (logrados por genes traídos de la petunia transgénica).
- Mejoras en el potencial nutricional de los alimentos. Por ejemplo en el tomate, se ha logrado introducir unos genes del maíz que aumentan la biosíntesis de kaempferol en más de un 60%, principalmente en la pulpa. También se ha introducido un gen de Petunia que aumenta la biosíntesis de la quercetina en más de un 70 %, principalmente en la piel. También hay ejemplos de aplicación de la ingeniería en papas y en algunas forrajeras.
- Mejoras en el potencial farmacéutico de las plantas. Algunos flavonoides presentes en las leguminosas, llamados isoflavonoides, pueden actuar como fitoestrógenos, lo cual ha generado interés en el uso de estos compuestos para tratar desórdenes hormonales en humanos. La ingeniería genética se utiliza para buscar la biosíntesis de isoflavonoides en plantas de cultivo donde normalmente están ausentes. Se ha realizado un experimento inicial exitoso en Arabidopsis thaliana.
- Supresión de la fertilidad del polen. En semillas de híbridos como el maíz se ha utilizado la ingeniería genética para que el polen generado por estas plantas híbridas fuera estéril. Por ejemplo en el maíz se ha logrado crackeando dos genes CHS: C2 y Whp, lo que da como resultado polen estéril de color blanco, producto de la ausencia de flavonoides en él. También hay ejemplos en híbridos de petunias y de la planta del tabaco.
- Biosíntesis de flavonoides por bacterias genéticamente modificadas. Recientemente mediante la ingeniería genética se ha logrado cultivar bacterias capaces de sintetizar flavonoides de tipo flavanonas[50]
Uso de los flavonoides para experimentación en otras áreas
Los flavonoides o sus vías biosintéticas han sido también utilizados para experimentar en otras áreas de la ciencia, por ejemplo:
Uso de las vías biosintéticas de los flavonoides como herramienta. Los flavonoides han contribuido en forma directa o indirecta en el descubrimiento de muchos principios biológicos en los últimos 150 años.[2] Dos ejemplos bien conocidos son el uso que Mendel le dio a los colores de las flores y las semillas de Pisum sativum, entre otros caracteres, para desarrollar sus teorías acerca de los mecanismos de la herencia; y el estudio de la pigmentación de los granos de maíz de Barbara McClintock que llevó al descubrimiento de los elementos móviles en el ADN.
Más recientemente, los análisis de la pigmentación en el maíz y sus tejidos vegetativos identificaron el fenómeno epigenético conocido como paramutación, en el que las interacciones entre alelos resultan en cambios heredables en la expresión genética (Chandler et al. 2000[51] ). De manera similar, los efectos de la expresión de flavonoides transgénicos en la pigmentación de la flor de petunia dejó al descubierto el fenómeno de cosupresión (Que y Jorgensen 1998[52] Metzlaff et al. 2000,[53] ).
La vía de los flavonoides también fue un sujeto de interés para los estudios de evolución, en particular en la Ipomoea purpurea, que ofrece recursos genéticos únicos y una larga historia de análisis (Iida et al. 1999[12] Rausher et al. 1999,[13] Durbin et al. 2000.[14] ). Estos estudios apoyan la idea de que las enzimas de la biosíntesis de los flavonoides fueron derivadas de enzimas del metabolismo primario, y que la duplicación de genes ha permitido la adaptación de esas enzimas a funciones específicas.
Además, la vía de los flavonoides, y la vía del fenilpropanoide de la que sale, están sirviendo de modelos experimentales para entender la organización intracelular del metabolismo, con unos trabajos recientes en alfalfa y Arabidopsis que proveen información nueva en la canalización de intermediarios (channeling of intermediates) y en el asemblaje de complejos multienzimáticos (ver review en Winkel-Shirley 2001[1] ).
Uso de los flavonoides en Botánica Sistemática. La Botánica Sistemática es la ciencia que se ocupa de establecer relaciones de parentesco entre las plantas a partir de sus características morfológicas, anatómicas, fisiológicas, su estructura del ADN, etc. La Botánica Sistemática asume que mientras más parecidas son dos plantas entre sí, más probable es que estén cercanamente emparentadas. Por eso mientras más características a analizar haya, más precisa va a ser la determinación del parentesco.Los flavonoides son extensamente utilizados en Botánica Sistemática, probablemente porque son fáciles de extraer e identificar[11] Debido a que son muy variables, son más útiles en determinar relaciones entre especies cercanamente emparentadas (o incluso en estudios de variación entre poblaciones de la misma especie), pero también son ocasionalmente útiles para determinar relaciones filogenéticas a niveles más altos (Bate-Smith 1968,[54] Crawford 1978,[55] Gornall et al. 1979,[56] Harborne y Turner 1984,[57] ). Finalmente, la diversidad en la estructura química de los flavonoides ha demostrado ser útil en estudios de hibridación entre especies (ver Alston y Turner 1963[58] Smith y Levin 1963,[59] Crawford y Giannasi 1982[60] ).
El papel que jugaron los flavonoides en establecer relaciones filogenéticas es indudable. Por ejemplo, la presencia de ciertos 5-desoxiflavonoides en las plantas del género Amphypteryngium (que usualmente había sido ubicado en su propia familia, Julianaceae, por ejemplo en Cronquist 1981) apoya su ubicación en las anacardiáceas.
Referencias
- ↑ a b c Winkel-Shirley, B. 2001. "Flavonoid Biosynthesis. A Colorful Model for Genetics, Biochemistry, Cell Biology, and Biotechnology". Plant Physiology 126: 485-493.
- ↑ a b c Winkel-Shirley, B. 2001b. "It takes a garden. How work on diverse plant species has contributed to an understanding of flavonoid metabolism". Plant Physiology 127: 1399-1404. (pdf aquí)
- ↑ Singleton VL. "Flavonoids". En: Childester CO, Mrak EM, Stewart GF (editores). Advances in Food Research. Academic Press, Nueva York. 149-242.
- ↑ Flavonoides (isoflavonoides y neoflavonoides). IUPAC Compendium of Chemical Terminology. (pdf aquí.
- ↑ Martínez-Flórez S., J. Gonález-Gallego, J. M. Culebras y M. J. Tuñón. 2002. "Los flavonoides: propiedades y acciones antioxidantes". Nutr Hosp 17: 271-278. (pdf aquí)
- ↑ Williams, CA, Grayer, RJ. 2004. "Anthocyanins and other flavonoids". Nat. Prod. Rep. 21: 539-573. (
- ↑ Major Types Of Chemical Compounds In Plants & Animals: Part II Flavonoids
- ↑ Trabajo práctico sobre la flor Trabajo Práctico sobre la flor
- ↑ a b c d e J. Palazón, R.M. Cusidó y C. Morales Metabolismo y significación biológica de los polifenoles del vino, Grupo de Biotecnología Vegetal, Facultad de Farmacia, Universidad de Barcelona.
- ↑ Mónica Noel Sánchez González y John P.N. Rosazza, Conversión de Chalconas a Auronas por Aspergillus alliaceus UI315 Resumen
- ↑ a b Judd, W. S. Campbell, C. S. Kellogg, E. A. Stevens, P.F. Donoghue, M. J. 2002. "Secondary Plant Compounds" en: Plant systematics: a phylogenetic approach, Second Edition. Sinauer Axxoc, USA. Capítulo 4.
- ↑ a b Iida S, A Hoshino, Y Johzuka-Hisatomi, Y Habu y Y Inagaki. 1999. "loricultural Traits and Transposable Elements in the Japanese and Common Morning Gloriesa". Annals of the New York Academy of Sciences 870: 265-274 (resumen y pdf aquí).
- ↑ a b Rausher MD, RE Miller, P Tiffin. 1999. "Patterns of evolutionary rate variation among genes of the anthocyanin biosynthetic pathway". Molecular Biology and Evolution 16: 266-274 (resumen y pdf aquí).
- ↑ a b Durbin ML, B McCaig, MT Clegg. 2000. "Molecular evolution of the chalcone synthase multigene family in the morning glory genome". Plant Molecular Biology 42: 79-92. (resumen aquí)
- ↑ Winkey-Shirley, B. 2001. "It takes a Garden. How work on diverse plant species has contributed to an understanding of flavonoid metabolism". Plant Physiology 127: 1399-1404.
- ↑ Kimura Y., T. Aoki, S. Ayae. 2001. "Chalcone isomerase isozimes with different substrate specifities towards 6'-hydroxy and 6'-deoxychalcones in cultured cells of Glycyrrhiza echinata, a leguminous plant producing 5-deoxyflavonoids". Plant Cell Physiol 42: 1169-1173.
- ↑ Brown D. E., A. M. Rashotte, A. S. Murphy, J. Normanly, B. W. Tague, W. A. Peer, L. Taiz, G. K. Muday. 2001. "Flavonoids act as negative regulators of auxin transport in vivo in Arabidopsis thaliana". Plant Physiol 126: 524-535.
- ↑ a b Principios activos de las plantas medicinales: los flavonoides Botanical Online
- ↑ Graf BA, Milbury PE, Blumberg, JB: "Flavonols, flavones, flavanones, and human health: epidemiological evidence." J Med Food 8: 281–290 (resumen aquí).
- ↑ Noticia: Curan el cáncer de hígado en un experimento con consumo de quercetina (Noticia de 20 minutos)
- ↑ M. G. Hertog et al. 1995. "Flavonoid intake and long-term risk of coronary heart disease and cancer in the seven countries study". Archives of Internal Medicine Vol. 155 No. 4. (resumen aquí, ver también la lista de artículos que lo citan)
- ↑ a b Yochum, L. et al. 1999. "Dietary Flavonoid Intake and Risk of Cardiovascular Disease in Postmenopausal Women." American Journal of Epidemiology 149:10 (resumen aquí)
- ↑ Balz Frei. 5 de marzo del 2007. "Studies force new view on biology of flavonoids". EurekAlert! (texto online aquí).
- ↑ a b Crozier, A. et al. 1997. "Quantitative Analysis of the Flavonoid Content of Commercial Tomatoes, Onions, Lettuce, and Celery" J. Agric. Food Chem., 45 (3), 590 -595. (resumen aquí)
- ↑ Kreuzaler F, Ragg H, Heller W, Tesch R, Witt I, Hammer D, Hahlbrock K. "Flavanone synthase from Petroselinum hortense. Molecular weight, subunit composition, size of messenger RNA, and absence of pantetheinyl residue." Eur J Biochem. 1979 99(1): 89–96. (resumen aquí)
- ↑ Moustafa, E. y Wong, E. 1967. "Purification and properties of chalcone-flavonone isomerase from soya bean seed." Phytochemistry 6: 625-632.
- ↑ Dixon RA, Dey PM, Whitehead IM. 1982. "Purification and properties of chalcone isomerase from cell suspension cultures of Phaseolus vulgaris". Biochim. Biophys. Acta 715: 25-33.
- ↑ Forkmann, G., Heller, W. y Grisebach, H. "Anthocyanin biosynthesis in flowers of Matthiola incana flavanone 3- and flavonoid 3'-hydroxylases." Z. Naturforsch. 35: 691-695
- ↑ Spribille, R. y Forkmann, G. 1984. "Conversion of dihydroflavonols to flavonols with enzyme extracts from flower buds of Matthiola incana". Z. Naturforsch. 39: 714-719.
- ↑ Britsch L y Grisebach H. 1986. "Purification and characterization of (2S)-flavanone 3-hydroxylase from Petunia hybrida". European Journal of Biochemistry 156: 569-577
- ↑ Stich K, Eidenberger T, Wurst F, Forkmann G. 1992. "Enzymatic conversion of dihydroflavonols to flavan-3,4-diols using flower extracts of Dianthus caryophyllus L.(Carnation)". Planta 187: 103–108.
- ↑ Federoff, N. V., Furtek, D. B. y Nelson, O. E. "Cloning of the Bronze Locus in Maize by a Simple and Generalizable Procedure Using the Transposable Controlling Element Activator (Ac)" Proc. natn. Acad. Sci. U.S.A. 81: 3825−3829. (resumen y pdf aquí)
- ↑ Devic M, J Guilleminot, I Debeaujon, N Bechtold, E Bensaude, M Koornneef, G Pelletier, M Delseny. 1999. "The BANYULS gene encodes a DFR-like protein and is a marker of early seed coat development" The Plant Journal 19 (4), 387–398. (resumen y pdf aquí)
- ↑ X Dong, EL Braun, E Grotewold. 2001. "Functional Conservation of Plant Secondary Metabolic Enzymes Revealed by Complementation of Arabidopsis Flavonoid Mutants with Maize Genes". Plant Physiology 127: 46-57 (resumen y pdf aquí).
- ↑ review en Dixon y Steele 1999
- ↑ Shirley, BW. 1996. "Flavonoid biosynthesis:'new' functions for an 'old' pathway" Trends Plant Sci 1:377-382.
- ↑ Rolfe BG. 1988. "Flavones and isoflavones as inducing substances of legume nodulation." Biofactors 1: 3-10 (resumen aquí)
- ↑ R Zerback, K Dressler, D Hess. 1989. "Flavonoid compounds from pollen and stigma of Petunia hybrida: inducers of the vir region of the Agrobacterium tumefaciens Ti plasmid". Plant Sci 62: 83-91.
- ↑ Kelly CK. 1990. " Plant Foraging: A Marginal Value Model and Coiling Response in Cuscuta Subinclusa". Ecology 71: 1916-1925 (resumen aquí).
- ↑ Albrecht H, JI Yoder, DA Phillips. 1999. "Flavonoids Promote Haustoria Formation in the Root Parasite Triphysaria versicolor" Plant Physiology 119: 585-591. (resumen y pdf aquí)
- ↑ Westwood JH. "Characterization of the Orobanche–Arabidopsis system for studying parasite–host interactions" Weed Science 48: 742-748 (resumen aquí).
- ↑ Li J, TM Ou-Lee, R Raba, RG Amundson, RL Last. 1993. "Arabidopsis Flavonoid Mutants Are Hypersensitive to UV-B Irradiation". The Plant Cell 5: 171-179 (resumen y pdf aquí)
- ↑ Debeaujon I, M Koornneef. 2000. "Gibberellin Requirement for Arabidopsis Seed Germination Is Determined Both by Testa Characteristics and Embryonic Abscisic Acid". Plant Physiology 122: 415-424 (resumen y pdf aquí).
- ↑ Jacobs M, PH Rubery. "Naturally Occurring Auxin Transport Regulators". Science 241: 346-349 (resumen aquí).
- ↑ Brown DE, AM Rashotte, AS Murphy, J Normanly, BW Tague, WA Peer, L Taiz y GK Muday. 2001. Plant Physiology 126: 524-535 (resumen y pdf aquí)
- ↑ Nesi N., C. Jond, I. Debeaujon, M. Caboche, L. Lepiniec. 2001. "The Arabidopsis TT2 gene encodes an R2R3 MYB domain protein that acts as a key determinant for accumulation in developing seed". Plant Cell 13: 2099-2114.
- ↑ Meyer P, Heidmann I, Forkmann G, Saedler H. 1987. "A new Petunia flower colour generated by transformation of a mutant with a maize gene". Nature 330: 667-668.
- ↑ Dixon R, Steele C. 1999. "Flavonoids and isoflavonoids - a gold mine for metabolic engineering". Trends Plant Sci 4: 394-400.
- ↑ Forkmann, G., S. Martens. 2001. "Metabolic engineering and applications of flavonoids". Current Opinion in Biotechnology 12: 155–160 (pdf aquí).
- ↑ Hwang E. I. , M. Kaneko , Y. Ohnishi , S. Horinouchi. 2003. "Production of plant-specific flavanones by Escherichia coli containing an artificial gene cluster." Appl Environ Microbiol. 69 (5): 2699-706
- ↑ Chandler VL, WB Eggleston, JE Dorweiler. 2000. "Paramutation in maize". Plant Molecular Biology 43: 121–145. (resumen aquí)
- ↑ Que Q, RA Jorgensen. 1998. "Homology-based control of gene expression patterns in transgenic petunia flowers". Developmental Genetics 22: 100-109 (resumen aquí).
- ↑ Metzlaff M, M O'dell, R Hellens, RB Flavell. 2000. "Developmentally and transgene regulated nuclear processing of primary transcripts of chalcone synthase A in petunia" The Plant Journal 23: 63–72. (resumen aquí).
- ↑ Bate-Smith, E. C. 1968. "The phenolic constituents of plants and their taxonomic significance" J. Linnean Soc. Bot. 60: 325-383.
- ↑ Crawford, D. J. 1978. "Flavonoid chemistry and angiosperm evolution." Bot. Rev. 44: 431-456.
- ↑ Gornall, R. J., B. A. Bohm y R. Dahlgre. 1979. "The distribution of flavonoids in the angiosperms". Bot. Notiser. 132: 1-30.
- ↑ Harborne, J. B. y B. L. Turner. 1984. Plant chemosystematics. Academic Press, Londres.
- ↑ Alston, R. E. y B. L. Turner. 1963. "Natural hybridization among four species of Baptisia (Leguminosae)". Am. J. Bot. 50: 159-173.
- ↑ Smith, D. M. y D. A. Levin. 1963. "A chromatographic study of reticulate evolution in the Appalachian Asplanium complex". Am. J. Bot. 50: 952-958.
- ↑ Crawford, D. J. y D. E. Giannasi. 1982. "Plant chemosystematics". BioScience. 32: 114-118, 123-124.
Bibliografía y enlaces
- Biotecnología en el Cultivo de Especies Ornamentales. Escandón, Alejandro S., Biotecnología y Mejoramiento Vegetal pp. 255-266
- Flavonoides, isoflavonoides y salud. María Rosario de Felipe y José Manuel Pozuelo, Centro de Ciencias Medioambientales (CSIC), Schironia Nº 3- Julio de 2004
- AN9, a Petunia Glutathione S-Transferase Required for Anthocyanin Sequestration, Is a Flavonoid-Binding Protein. Lukas A. Mueller, Christopher D. Goodman, Rebecca A. Silady, and Virginia Walbot, Plant Physiology, August 2000, Vol. 123, pp. 1561–1570,
- General Plant Metabolism. Organised by N. Smirnoff for the Plant Metabolism Group, Abstracts / Comparative Biochemistry and Physiology Part A 132 (2002) S173–S180
- The characterisation of New Zealand Podocarpus hybrids using flavonoid markers. ROSEMARY F. WEBBY, KENNETH R. MARKHAM, New Zealand Journal of Botany, 1987, Vol. 25: 355-366
- The Unique Occurrence of the Flavone Aglycone Tricetin in Myrtaceae Pollen. Maria G. Camposa, Rosemary F. Webbyb and Kenneth R. Markhamb, 0939Ð5075/2002/0900Ð0944 $ 06.00 ” 2002 Verlag der Zeitschrift für Naturforschung, Tübingen
- Estudio del efecto solvatocrómico en derivados fenólicos naturales. Romero Ale, E; Olives, AI; Martín, L; Martín, MA;* Del Castillo, B.; Agnese, AM.; Ortega, MG; Núñez-Montoya, S; Cabrera JL*, Ars Pharmaceutica, 43:1-2; 57-71, 2002
- Curso Popular de Cata de Vinos. 1997, Manuel Ruiz Hernández, Miguel Martínez Garoña
- Arabidopsis ICX1 Is a Negative Regulator of Several Pathways Regulating Flavonoid Biosynthesis Genes. Helena K. Wade, Awinder K. Sohal2, and Gareth I. Jenkins, Plant Physiology, February 2003, Vol. 131, pp. 707–715,
- Section E SNIF part 2: Summary information format for products containing genetically modified higher plants (GMHPs). Florigene Moonlite (123.2.28)
Enlaces relacionados no bibliográficos
- Anthocyanins and other flavonoids. J. B. Harborne and C. A. Williams, Plant Science Laboratories, University of Reading, Reading, UK, RG6 6AS (Muy bueno para buscar qué flavonoides se presentan en determinadas plantas).
- Horse Chestnut (Aesculus hippocastanum). Mary McLellan, RN, BSN, Longwood Herbal Task Force, Revised June 15, 2000
Categoría:- Flavonoides





Wikimedia foundation. 2010.