- Fabaceae
-
 Fabáceas (Leguminosas)
Fabáceas (Leguminosas)
Vicia sativaClasificación científica Reino: Plantae División: Angiospermae Clase: Eudicotyledoneae Orden: Fabales Familia: Fabaceae Lindley (Leguminosae Jussieu, nom. cons.).[1] Diversidad 730 géneros y 19.400 especies (ver Anexo:Géneros de Fabaceae) Subfamilias - Faboideae Rudd (Papilionoideae Juss., es el nombre alternativo)
- Caesalpinioideae DC.
- Mimosoideae DC.
Sinonimia Las fabáceas (Fabaceae) o leguminosas (Leguminosae)[2] son una familia del orden de las fabales. Reúne árboles, arbustos y hierbas perennes o anuales, fácilmente reconocibles por su fruto tipo legumbre y sus hojas compuestas y estipuladas. Es una familia de distribución cosmopolita con aproximadamente 730 géneros y unas 19.400 especies, lo que la convierte en la tercera familia con mayor riqueza de especies después de las compuestas (Asteraceae) y las orquídeas (Orchidaceae).[3] [4] Esta riqueza de especies se halla particularmente concentrada en las ramas de las mimosóideas y las fabóideas, ya que contienen cerca del 9,4% de la totalidad de las especies de las eudicotiledóneas.[5] Se ha estimado que alrededor del 16% de todas las especies arbóreas en los bosques lluviosos neotropicales son miembros de esta familia. Asimismo, las fabáceas son la familia más representada en los bosques tropicales lluviosos y en los bosques secos de América y África.[6]
Independientemente de los desacuerdos que hasta hace poco tiempo existieron en torno a si las fabáceas deberían ser tratadas como una sola familia compuesta de tres subfamilias o como tres familias separadas, existe una gran cantidad de información y evidencias tanto moleculares como morfológicas que sustentan que las leguminosas son una única familia monofilética.[7] Este punto de vista se ha reforzado no solo por el grado de interrelación que exhiben diferentes grupos dentro de la familia comparados con aquel hallado entre las leguminosas y sus parientes más cercanos, sino también por todos los recientes análisis filogenéticos basados en secuencias del ADN.[8] [9] [10] Tales estudios confirman que las leguminosas son un grupo monofilético y que está estrechamente relacionado con las familias Polygalaceae, Surianaceae y Quillajaceae junto a las que conforman el orden Fabales.[11]
Junto con los cereales y con algunas frutas y raíces tropicales, varias leguminosas han sido la base de la alimentación humana durante milenios, siendo su utilización un compañero inseparable de la evolución del hombre.[12]
Contenido
Descripción
Hábito
Las leguminosas presentan una gran variedad de hábitos de crecimiento, pudiendo ser desde árboles, arbustos o hierbas, hasta enredaderas herbáceas o lianas. Las hierbas, a su vez, pueden ser anuales, bienales o perennes, sin agregaciones de hojas basales o terminales. Son plantas erguidas, epífitas o enredaderas. En este último caso se sostienen mediante los tallos que se retuercen sobre el soporte o bien por medio de zarcillos foliares o caulinares. Pueden ser heliofíticas, mesofíticas o xerofíticas.[1] [3]
Hojas
Las hojas son casi siempre alternas y con estípulas, persistentes o caedizas, generalmente compuestas, pinnadas o bipinnadas, digitadas o trifoliadas, a veces aparentemente simples —es decir, unifoliadas o ausentes y, en ese caso, los tallos se hallan transformados en filodios— o pinnadas y con zarcillos en el ápice. A menudo las hojas se hallan reducidas o son precozmente caducas o nulas en las especies áfilas o subáfilas. El pecíolo, y muchas veces los peciolulos, tienen la base engrosada, "ganglionar", que permite movimientos —las denominadas posiciones de "sueño" y de "vigilia"—. Es frecuente la presencia de espinas por transformación del raquis de las hojas, de las estípulas o del tallo.[1] [12] [3]
Raíz
Las raíces presentan un predominio del sistema primario, es decir, de aquél que proviene de la radícula del embrión. Las raíces de las leguminosas son a menudo profundas y casi siempre exhiben nódulos poblados de bacterias del género Rhizobium que asimilan el nitrógeno atmosférico.[12]
Flor
Las flores pueden ser desde pequeñas o grandes, actinomorfas —en el caso de las mimosóideas— a leves o profundamente cigomorfas —como ocurre en las papilionóideas y en la mayor parte de las cesalpinióideas—. Las irregularidades en la simetría floral en estos casos involucran al perianto y al androceo. El receptáculo de la flor desarrolla un «ginóforo» —frecuentemente fusionado al hipanto en las cesalpinióideas— con forma de cúpula. El hipanto puede estar presente o ausente, en este último caso está reemplazado por el tubo del cáliz, como es el caso en la mayoría de las fabóideas. El perianto tiene casi siempre un cáliz y una corola diferenciadas. No obstante, la corola puede estar ausente, en cuyo caso el perianto se dice «sepalino» (similar a sépalos), como ocurre en decenas de géneros de cesalpinióideas y algunas especies de las tribus Swartzieae y Amorphieae. El cáliz presenta cinco sépalos —raramente tres o seis— dispuestos en un sólo ciclo, los cuales pueden estar total o parcialmente unidos entre sí. El cáliz, además, puede ser o no persistente, raramente es acrescente (es decir que continúa en el fruto), imbricado o valvado. La corola está compuesta por cinco pétalos libres —comúnmente menos de cinco o ausentes en Swartzieae, Amorphieae y en las cesalpinióideas, o tres a cuatro en las mimosóideas— o parcialmente unidos, y presenta, en general, una morfología característica. Así, la corola papilionácea o amariposada está integrada por un pétalo superior muy desarrollado, conocido como «estandarte» o «vexilo», dos pétalos laterales o «alas» y dos piezas inferiores a menudo conniventes que constituyen una estructura simpétala denominada «carena» o «quilla». Esta arquitectura es muy similar a la de las flores de las cesalpinóideas pero, a diferencia de lo que ocurre en éstas, con prefloración vexilar o descendente, es decir, con el estandarte recubriendo el resto de las piezas corolinas dentro del botón floral.[1] El gineceo es de ovario súpero, con un solo carpelo, con desarrollo muy variable y tendencia a la reducción en el número de óvulos.[12] [3] [4]
Inflorescencia
Las flores son solitarias o pueden disponerse en diversos tipos de inflorescencias: racimos terminales o axilares, a veces se modifican hasta parecer cabezuelas y, en otras ocasiones, umbelas.[12] [3]
Fruto
El fruto de las leguminosas, técnicamente denominado legumbre, deriva de un ovario compuesto por un sólo carpelo el cual —en la madurez— se abre longitudinalmente en dos valvas, lo que indica que su dehiscencia ocurre por la nervadura media y por la unión carpelar. No obstante, existe una inmensa variedad de formas y tamaños de frutos en la familia. De hecho, hay especies con frutos con tendencia a la indehiscencia —es decir que no se abren en la madurez— y en ocasiones el cáliz se transforma en una estructura de dispersión. Uno de los frutos más notables de la familia tal vez sea el del maní (género Arachis) ya que las flores, tras la polinización, se hunden en el suelo y el fruto (una legumbre indehiscente) se desarrolla subterráneamente. No obstante, las legumbres más grandes las produce una liana tropical perteneciente al género Entada. Las legumbres más largas de la especie centroamericana Entada gigas (llamada "escalera de mono" en su tierra natal) llegan a medir hasta 1,5 m de longitud. Otras especies han desarrollado curiosos mecanismos de dispersión. Así, la especie sudamericana Tipuana tipu ha modificado parte de la pared del fruto para convertirla en un ala, la cual le permite trasladarse con el viento rotando como las aspas de un helicóptero. Ese tipo de fruto se denomina sámara.[1] [3] [12] [13]
 Follaje y frutos de la especie Tipuana tipu.
Follaje y frutos de la especie Tipuana tipu.
Semillas
Las semillas no tienen endosperma. En cambio, acumulan en los cotiledones sobre todo almidón y proteínas, a veces aceites, o aceites y proteínas. Lo más característico es la presencia de grandes cotiledones ricos en reservas, a menudo oleaginosas (como por ejemplo en la soja o el cacahuate).[12] [3]
Fisiología y bioquímica
Las leguminosas raramente son cianogenéticas y, en ese caso, los compuestos cianogenéticos derivan de la tirosina, la fenilalanina o de la leucina. Comúnmente presentan alcaloides. Las protoantocianidinas pueden estar presentes y, en ese caso, son la cianidina, la delfinidina o ambas a la vez. Frecuentemente presentan flavonoides tales como kaempferol, quercitina y miricetina. El ácido elágico se halla consistentemente ausente en todos los géneros y especies analizadas de las tres subfamilias. Los azúcares se transportan dentro de la planta en forma de sacarosa. La fisiología C3 se ha demostrado e informado en una gran cantidad de géneros de las tres subfamilias.[1]
Ecología
Distribución y hábitat
Es una familia de distribución cosmopolita. Los árboles son más frecuentes en las regiones tropicales, mientras que las hierbas y los arbustos dominan en las extratropicales.[1]
Fijación biológica del nitrógeno
 Raíces de Vicia en las que se pueden observar los nódulos radiculares de color blanco.
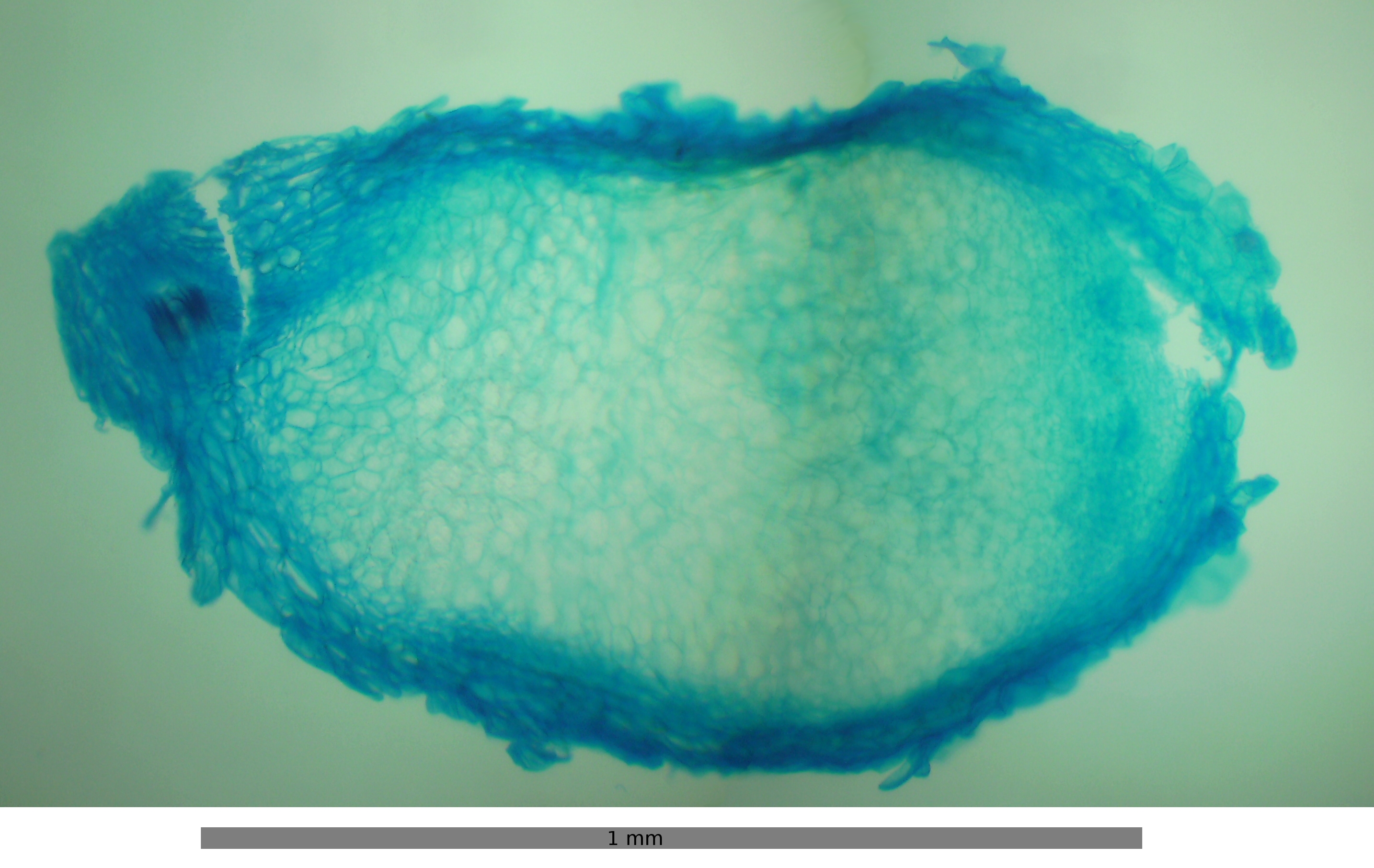
Raíces de Vicia en las que se pueden observar los nódulos radiculares de color blanco.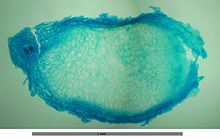 Sección transversal de un nódulo radicular de Vicia observado al microscopio.
Sección transversal de un nódulo radicular de Vicia observado al microscopio.La fijación biológica del nitrógeno (FBN, diazotrofía) es un proceso muy antiguo que probablemente se originó en el Eon arqueano bajo las condiciones de ausencia de oxígeno de la atmósfera primitiva. Es exclusivo de Euryarchaeota y en 6 de los más de 50 phyla de Bacteria. Algunos de estos linajes coevolucionaron conjuntamente con las angiospermas estableciendo las bases moleculares de una relación de simbiosis mutualista. Los nódulos son las estructuras especializadas dentro de las cuales se lleva a cabo la FBN que algunas familias de angiospermas han desarrollado, principalmente en la corteza radicular y excepcionalmente en el tallo como es el caso de Sesbania rostrata. Las fanerógamas que han coevolucionado con diazótrofos actinorrícicos o con los rizobios para establecer su relación simbiótica pertenecen a 11 familias que se agrupan dentro del clado Rosidae de la filogenia molecular del gen rbcL que codifica parte de la enzima RuBisCO en el cloroplasto. Este agrupamiento indica que probablemente la predisposición a la formación de nódulos surgió una sola vez en las angiospermas y podría ser considerado como un carácter ancestral que se ha conservado o perdido en ciertos linajes. Sin embargo, la distribución tan dispersa de familias y géneros nodulantes dentro de este linaje, indica orígenes múltiples de la nodulación. De las 10 familias nodulantes de Rosidae, 8 son noduladas por actinomicetos (Betulaceae, Casuarinaceae, Coriariaceae, Datiscaceae, Elaeagnaceae, Myricaceae, Rhamnaceae y Rosaceae), y las dos familias restantes Ulmaceae y Fabaceae son noduladas por rizobios.[14]
«Rizobios» es el nombre que recibe el grupo de géneros de alfa-proteobacterias (familia Rhizobiaceae) que incluyen todas las especies que fijan nitrógeno y que producen nódulos con las leguminosas, tales como Allorhizobium, Azorhizobium, Bradyrhizobium, Mesorhizobium, Phyllobacterium, Rhizobium y Sinorhizobium, así como el fitopatógeno Agrobacterium.[15] Los rizobios y sus leguminosas hospedantes se deben reconocer uno al otro para que la nodulación se inicie. Diferentes especies de rizobios son específicas de sus especies hospedantes, pero pueden infectar -frecuentemente- a más de una especie. Asimismo, una sola especie de planta puede ser infectada por más de una especie de bacteria. Acacia senegal, por ejemplo, puede ser nodulada por siete especies de rizobios pertenecientes a tres géneros distintos. Los caracteres más distintivos que permiten distinguir a los géneros de rizobios son su rapidez de crecimiento y el tipo de nódulo radical que forman con su hospedante.[15] A su vez, los nódulos radicales se clasifican en indeterminados, que son cilíndricos y frecuentemente ramificados; y determinados, los cuales son esféricos y con lenticelas prominentes. Los nódulos indeterminados son característicos de las leguminosas de climas templados, mientras que los determinados son usuales en las especies de climas tropicales a subtropicales.[15] La nodulación prosperó ampliamente dentro de las leguminosas; se encuentra presente en la mayoría de sus miembros que se especializaron en la asociación exclusiva con rizobios, los que a su vez también establecieron una simbiosis exclusiva con las leguminosas (con la sola excepción de Parasponia, el único género de los 18 que se incluyen en Ulmaceae que es capaz de nodular). La nodulación está presente en todas las subfamilias de leguminosas pero es menos frecuente en la subfamilia Caesalpinioideae. La subfamilia Papilionoideae es la que presenta todos los tipos de nódulos: indeterminado (conserva el meristemo), determinado (no conserva el meristemo) y el tipo de Aeschynomene. Los dos últimos, sólo están presentes en algunas líneas de la subfamilia Papilionoideae, por lo que son consideradas como los tipos de nódulos más modernos y especializados. Aunque la nodulación es abundante en las dos subfamilias monofiléticas Papilionoideae y Mimosoideae, también existen ejemplos de especies no nodulantes. La presencia y ausencia de especies nodulantes dentro de las tres subfamilias indica que la nodulación surgió varias veces en el transcurso de la evolución de las leguminosas y que se ha perdido en algunos linajes. Por ejemplo, dentro del género Acacia, miembro de Mimosoideae, A. pentagona no tiene la capacidad de nodular, mientras que otras especies del mismo género lo hacen de modo promiscuo, como es el caso de la ya citada Acacia senegal que es nodulada por rizobios de rápido y lento crecimiento.[14]
Evolución, filogenia y sistemática
Evolución
El orden Fabales contiene cerca del 9,6% de la diversidad de especies de las eudicotiledóneas y la mayor parte de esa riqueza se concentra en una de las cuatro familias que lo componen: Fabaceae. El origen de este clado, compuesto además por las familias Polygalaceae, Surianaceae y Quillajaceae, está datado en 94 a 89 millones de años, comenzando su diversificación hace unos 79 a 74 millones de años.[4] De hecho, aparentemente las leguminosas se han diversificado durante el terciario temprano para convertirse en un integrante ubicuo de las biotas terrestres modernas, al igual que muchas otras familias de Angiospermas.[16] [17]
El registro fósil de las leguminosas es abundante y diverso, particularmente en el período Terciario, con fósiles de flores, frutos, folíolos, madera y polen conocidos de numerosas localidades.[18] [19] [20] [21] [22] Los primeros fósiles que se pueden asignar definitivamente a las leguminosas aparecen en el Paleoceno tardío (hace cerca de 56 millones de años).[23] [24] Representantes de las 3 subfamilias tradicionalmente reconocidas de las leguminosas —cesalpinióideas, papilionóideas y mimosóideas— así como también de grandes clados dentro de esas subfamilias —como el de las genistóides— se han hallado en el registro fósil en períodos un poco más tardíos, comenzando entre 55 a 50 millones de años atrás.[16] De hecho, el hallazgo de diversos grupos de taxones que representan los principales linajes de las leguminosas durante el Eoceno medio y el Eoceno tardío sugiere que la mayoría de los grupos modernos de fabáceas ya estaban presentes y que en ese período ocurrió una extensa diversificación.[16] Las fabáceas, entonces, iniciaron su diversificación hace aproximadamente 60 millones de años y los clados más importantes se han separado hace unos 50 millones de años.[25] La edad de los principales clados de las cesalpinióideas se ha estimado entre 56 a 34 millones de años y la del grupo basal de las mimosóideas en 44 ± 2,6 millones de años.[26] [27] La diferenciación entre mimosóideas y fabóideas está datada en 59 a 34 millones de años antes del presente y la edad del grupo basal de las fabóideas en 58,6 ± 0,2 millones de años atrás.[28] Dentro de las fabóideas, la divergencia de algunos grupos ha podido ser datada. Así, Astragalus se ha separado de Oxytropsis hace unos 16 a 12 millones de años; a pesar que la diversificación dentro de cada género fue relativamente reciente. Por ejemplo, la radiación de las especies aneuploides de Neoastragalus comenzó hace 4 millones de años. Inga, otro género de papilionóideas con aproximadamente 350 especies, parece haber divergido en los últimos 2 millones de años.[29] [30] [31] [32]
Filogenia y taxonomía
La filogenia de las leguminosas ha sido el objeto de estudio de numerosos grupos de investigación de todo el mundo, los cuales han utilizado la morfología o los datos del ADN (el intrón plastídico trnL, los genes plastídicos rbcL y matK, o los espaciadores ribosomales ITS) y el análisis cladístico para resolver las relaciones entre los diferentes linajes de esta familia. Así, se ha podido confirmar la monofilia de las subfamilias tradicionales Mimosoideae y Papilionoideae, anidadas conjuntamente con la subfamilia parafilética Caesalpinioideae. Todos los estudios, a través de diferentes aproximaciones, han sido consistentes, lo que brinda un gran apoyo a las relaciones entre los principales clados de la familia, como se muestra en el cladograma de más abajo.[4] El clado que más tempranamente se ramifica es la tribu Cercideae, seguido por dos clados compuestos de taxones antes dispuestos dentro de Caesalpinioideae (la tribu Detarieae y el género Duparquetia ).[33] [34] [35] [36] [37] [38]
Polygalaceae Surianaceae Fabales Quillajaceae Tribus Cercideae, Detarieae y el género Duparquetia Fabaceae Caesalpinioideae Mimosoideae Faboideae Sobre la base de las relaciones filogenéticas descritas, se analizarán a continuación cada uno de los grupos que componen a esta familia.
1a. Cercideae
Comprende árboles y lianas, las hojas son aparentemente simples, algunas veces bilobadas. El número cromosómico básico es x=7. Incluye de 4 a 12 géneros y unas 265 especies. El género más representado es Bauhinia con unas 250 especies de distribución pantropical. Cercis, otro de los géneros dentro de esta tribu, presenta flores que, en apariencia, son similares a las fabóideas. Otros de los géneros incluidos en este clado son Caesalpinia y Delonix.[4] Sinonimia: Bauhiniaceae Martynov.
1b. Duparquetia
Este género presenta características particulares en cuanto a su morfología y desarrollo floral: un cáliz de cuatro sépalos y un androceo de cuatro estambres lo que las separa netamente de las restantes fabáceas. El clado está representado por un género monotípico, cuya única especie se denomina Duparquetia orchidacea y se distribuye por África tropical.[4]
1c. Detarieae
Presentan el envés (la cara de abajo) de las hojas cubiertas por glándulas con forma de cráter y las estípulas y bractéolas deciduas, caedizas. Los miembros de la tribu muestran una reducción en el número de pétalos o de estambres. Producen, además, un tipo de resina muy particular, la cual contiene diterpenos bicíclicos.[4] La tribu incluye muchos árboles tropicales entre ellos algunos que se utilizan para la producción de madera. Incluye unos 80 géneros con un número cromosómico básico de x=12. Se distribuyen en África y Sudamérica.[39]
2. Cesalpinióideas
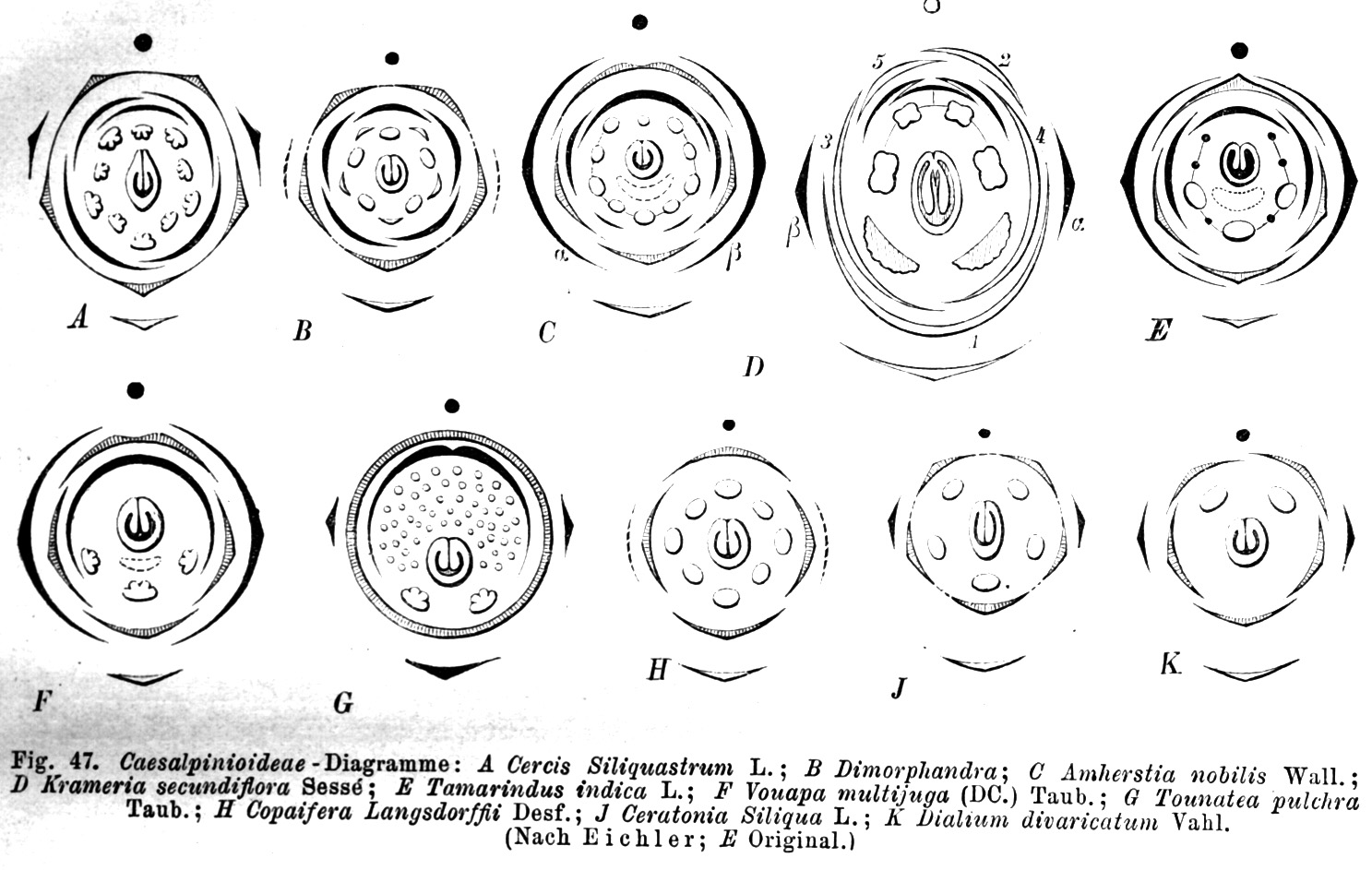
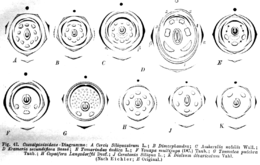 Diagramas florales de varias especies de cesalpinióideas que muestran la gran variabilidad de la morfología floral de esta subfamilia. A. Cercis siliquastrum. B. Dimorphandra. C. Amherstia nobilis. D. Krameria secundiflora. E. Tamarindus indica. F. Vouapa multijuga. G. Tounatea pulchra. H. Copaifera langsdorfii. J. Ceratonia siliqua. K. Dialium divaricatum.
Diagramas florales de varias especies de cesalpinióideas que muestran la gran variabilidad de la morfología floral de esta subfamilia. A. Cercis siliquastrum. B. Dimorphandra. C. Amherstia nobilis. D. Krameria secundiflora. E. Tamarindus indica. F. Vouapa multijuga. G. Tounatea pulchra. H. Copaifera langsdorfii. J. Ceratonia siliqua. K. Dialium divaricatum.Son árboles, subarbustos o, a veces, hierbas o trepadoras. Las hojas son pinnadas o bipinnadas, en algunos casos (por ejemplo, en Gleditsia) ambos tipos foliares se hallan en el mismo árbol; finalmente, hay especies áfilas.
Las flores son cigomorfas, rara vez actinomorfas. La prefloración es imbricada, rara vez valvar; se dice "ascendente": el estandarte es interno, cubierto en sus bordes por los pétalos vecinos. La corola es variada, grande, mediana o pequeña, tiene pétalos libres (es dialipétala) por lo menos en la base, muy a menudo los pétalos son unguiculados. El androceo está formado por lo general por 10 estambres, libres o soldados en grupos, igualando la altura de los pétalos o cubiertos por ellos. El polen es libre y el tegumento seminal no presenta "línea fisural". El óvulo es anátropo. Las semillas tienen embrión recto, hilo apical o subapical, pequeño. La estructura floral es mucho más variada que en las mimosóideas. Las flores se disponen en racimos o panojas, en general de medianas a grandes, rara vez pequeñas, y en espiga densa. El fruto es dehiscente o indehiscente, muy variable. Habitan preferentemente en las regiones cálidas, y están muy bien representadas en las regiones tropicales de África y América, donde son especies a menudo gigantescas.[12] La subfamilia está compuesta por 160 géneros y aproximadamente 1.900 especies. Los géneros más representados son Senna (con 350 especies), Chamaechrista (265) y Caesalpinia (100).[4]
Forman parte de esta subfamilia plantas conocidas como cassias, cina-cina, pata de vaca, el algarrobo europeo, la acacia negra o Gleditsia, el guayacán y el chivato. Varias especies de cesalpinióideas se cultivan con fines decorativos, forestales, industriales o medicinales.[12]
Las Cesalpinióideas, de acuerdo a las últimas investigaciones filogenéticas, comprenden cuatro tribus: Cassieae, Caesalpinieae, Cercideae y Detarieae.[4]
3. Mimosóideas


Son árboles o arbustos, rara vez hierbas, de follaje grácil por sus hojas bipinnadas, raramente pinnadas o reducidas a filodios. En muchos casos son espinosos y de aspecto muy particular por las pequeñas flores agregadas en cabezuelas o espigas que, a su vez, suelen formar racimos espiciformes densos o grandes panojas.
Las flores son actinomorfas. La corola tiene prefloración valvar, a menudo es gamopétala y tubulosa. El cáliz es valvar o, en algunos grupos, imbricado, siempre gamosépalo. El androceo está formado por 4, 8, 10 o numerosos estambres, los cuales superan la altura de la corola. Los filamentos estaminales están libres o soldados entre sí. El polen es generalmente compuesto. El óvulo es anátropo. La semilla presenta el embrión recto y el tegumento presenta una "línea fisural" con forma de anillo o herradura. El hilo es apical o subapical, pequeño.[12]
Es un grupo muy bien representado por numerosos géneros y especies en regiones tropicales o subtropicales, de todos los continentes, pero están particularmente distribuidas en África y América. Comprenden 82 géneros y alrededor de 3.300 especies. Los géneros con mayor número de especies son Acacia sensu stricto (960, comprende solamente a las especies antes incluidas en el subgénero Phyllodinae), Mimosa (480), Inga (350), Calliandra (200), Vachellia (161, previamente conocido como Acacia subgenus Acacia), Senegalia (85, previamente incluidas dentro de Acacia subgenus Aculeiferum), Prosopis (45), Pithecellobium (40). Especies muy conocidas pertenecen a las mimosóideas, tales como las verdaderas acacias, las mimosas o sensitivas, los ingáes, timbóes y plumerillos, los aromos, tuscas, garabatos, espinillos y algarrobos. Abundan entre los grandes árboles de los bosques tropicales, dominan en muchas sabanas y son siempre frecuentes en las regiones semidesérticas cálidas. Muchas mimosóideas se cultivan como árboles para sombra, plantas ornamentales, de protección o con fines forestales.[12] La subfamilia comprende cinco tribus: Acacieae, Ingeae, Mimoseae, Mimozygantheae y Parkieae.
4. Fabóideas o papilionóideas
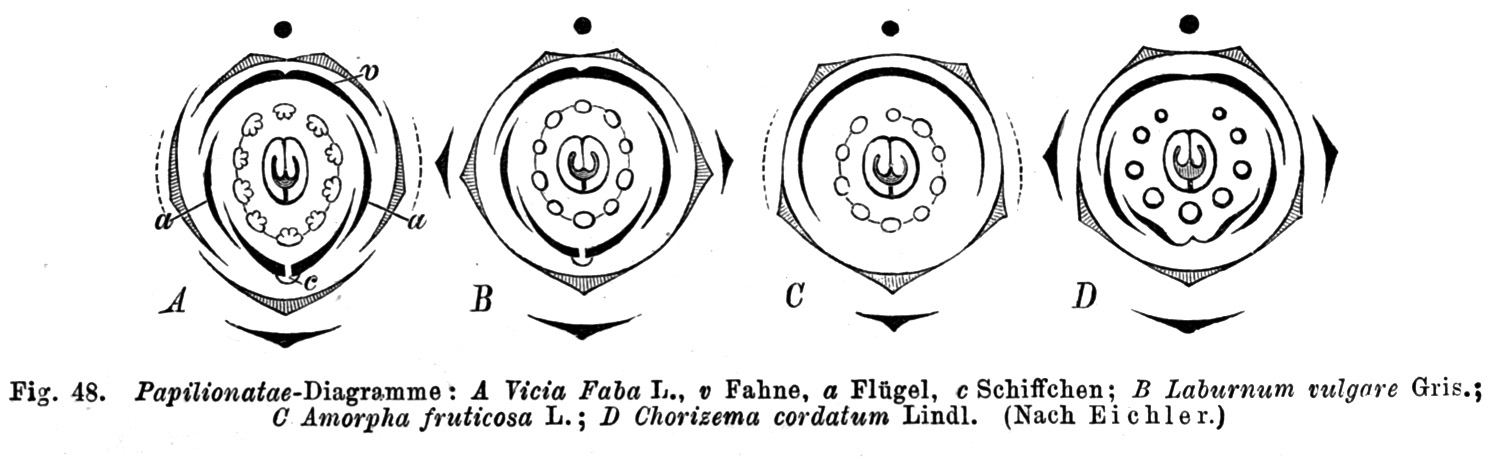
 Diagramas florales de algunas leguminosas fabóideas. A. Vicia faba. B. Laburnum vulgare. C. Amorpha fruticosa. D. Chorizema cordatum.
Diagramas florales de algunas leguminosas fabóideas. A. Vicia faba. B. Laburnum vulgare. C. Amorpha fruticosa. D. Chorizema cordatum.
Son árboles, arbustos, trepadoras leñosas o herbáceas o anuales. Las hojas son, por lo general, pinnadas o digitadas, frecuentemente trifoliadas, algunas veces unifolidas y hasta nulas, pero nunca bipinnadas. Las flores se disponen en racimos, panojas, capítulos o espigas. Las flores son cigomorfas, rara vez actinomorfas. La prefloración es imbricada, rara vez valvar; la prefloración de la corola se dice descendente: el estandarte es externo, cubriendo los pétalos vecinos. La corola es generalmente "amariposada" o "papilionoidea", con los dos pétalos inferiores conniventes o unidos en el ápice, formando la quilla, los dos laterales extendidos como alas y el superior erguido o reflejo, de mayor tamaño, llamado estandarte. EL cáliz está formado por 5 sépalos soldados entre sí y es campanulado o tubuloso. El androceo está formado por 10 estambres, rara vez menos, generalmente escondidos dentro de la quilla, y se hallan más o menos soldados entre sí; típicamente son diadelfos (9+1), pero también poliadelfos o libres. Los óvulos son campilótropos. Las semillas presentan un embrión curvado, raramente recto; el hilo es de tamaño mediano a grande.
Con 476 géneros y 13.860 especies, es la división más grande y diversa de la familia. Se extiende por todos los continentes excepto las regiones árticas e incluye la mayoría de las leguminosas más populares utilizadas como alimento o forraje, así como varias especies modelo para investigaciones genómicas.[40] Los géneros con mayor número de especies son Astragalus (2.400-3.270), Indigofera (700), Crotalaria (700), Mirbelia sensu lato (450), Tephrosia (350), Desmodium (300), Aspalanthus (300), Oxytropis (300), Adesmia (240), Trifolium (240), Rhynchosia (230), Lupinus (200), Aeschyomene (160), Hedysarum (160), Lathyrus (160), Vicia (160), Dalea (150), Eriosema (150), Lotononis (150), Millettia (150), Vigna (150), Swartzia (140), Daviesia (135), Lonchocarpus (135), Machaerium (130), Onobrychis (130), Ormosia (130), Lotus (inc. Coronilla, (125)), Lonchocarpus (120), Erythrina (110), Gastrolobium (110), Mucuna (105) y Pultenaea (100). Las especies leñosas son preferiblemente termófilas; muchas herbáceas son de regiones templadas y frías, si bien están también representadas en los trópicos. Comprende muchas plantas útiles y muy conocidas, tales como arveja, garbanzo, poroto o frijol, arvejilla, tréboles, haba, alfalfa, soja, entre otras.[12] [32] La subfamilia de las fabóideas se ha subdividido en las siguientes tribus, de acuerdo a las últimas investigaciones filogenéticas (APW): Abreae - Adesmieae - Aeschynomeneae - Amorpheae - Bossiaeeae - Brongniartieae - Carmichaelieae - Cicereae - Crotalarieae - Dalbergieae - Desmodieae - Dipteryxeae - Euchresteae - Fabeae - Galegeae - Genisteae - Hedysareae - Hypocalypteae - Indigofereae - Loteae - Millettieae - Mirbelieae - Phaseoleae - Podalyrieae - Psoraleeae - Robinieae - Sophoreae - Swartzieae - Thermopsideae - Trifolieae.
Importancia económica y cultural
Las leguminosas presentan una destacada importancia económica y cultural debido a su extraordinaria diversidad y abundancia de representantes en diversas formaciones vegetales y por sus aplicaciones: jardinería y ornamentación, alimento, obtención de compuestos con interés medicinal y en droguería, extracción de aceites y grasas.[13] [41] [42] [43]
Alimenticias
Durante miles de años, las legumbres han sido utilizadas como alimento por los seres humanos. La lenteja fue probablemente una de las primeras especies en ser domesticadas. Pueden consumirse frescas o secas, y pueden encontrarse en una gran variedad de colores, sabores y texturas. Se las conserva secas, enlatadas, o bien, congeladas. Todas las legumbres son muy similares en cuanto a sus características nutricionales. Son muy ricas en proteínas, carbohidratos y fibras, mientras que el contenido en lípidos es relativamente bajo y los ácidos grasos que lo componen son insaturados. Una gran excepción a esta regla es la soja, que tiene un alto contenido en lípidos. Las legumbres son una fuente importante de vitamina B. Las que se consumen frescas tienen vitamina C, pero los niveles declinan después de la cosecha y son virtualmente inexistentes en los granos secos. Las legumbres enlatadas, no obstante, mantienen aproximadamente la mitad de los valores de vitamina C que las legumbres frescas (excepto en el caso de las arvejas o lentejas que se secan antes de enlatarlas). Las arvejas congeladas pierden una cuarta parte de su contenido inicial de vitamina C. Las legumbres se consumen por su alto contenido de proteína. La composición de los granos de poroto o frijol, a modo de ejemplo, es de 21% de proteína, 46% de carbohidratos, 25% de fibra y 1,5% de lípidos. Contiene además, aproximadamente 7 mg de hierro y 180 mg de calcio por cada 100 gramos. Los granos de soja, en cambio, contienen 34% de proteína, 29% de carbohidratos y 18% de lípidos. El mayor contenido en proteínas y lípidos le otorgan a la soja una calidad nutricional superior al de otras legumbres.[44]
Entre los factores positivos de las leguminosas en la nutrición humana, cabe citar su alta concentración en proteínas con alto contenido en lisina, que hace a las leguminosas un complemento proteico excelente para los cereales. Son también una buena fuente de algunos minerales como calcio, hierro, zinc, fósforo, potasio y magnesio, así como de algunas vitaminas hidrosolubles, especialmente tiamina, riboflavina y niacina. En relación con la salud, la ingestión de algunas leguminosas producen una disminución del colesterol posiblemente debido a su alto contenido en fibra dietética y también pueden ayudar a reducir los niveles de glucosa sanguínea en los diabéticos. Entre los factores negativos se puede reseñar la deficiencia en aminoácidos azufrados y la presencia de ciertos factores antinutricionales que influyen en la digestibilidad proteica y de carbohidratos. Esto puede ser un factor limitante en las dietas de los vegetarianos y en países en vías de desarrollo donde la ingestión de proteína animal es baja o nula. Sin embargo, cuando la dieta es equilibrada o muy rica en proteína animal y energía, la ingestión de leguminosas es una ventaja.[45]
Los antinutrientes son sustancias que, en general, dificultan la asimilación de los nutrientes, y en algunos casos, pueden llegar a ser tóxicos o causar efectos fisiológicos poco deseables (como, por ejemplo, la flatulencia). Recientemente se ha visto que, en pequeñas cantidades, pueden ser también muy beneficiosos para la salud en la prevención de enfermedades como cáncer y enfermedades coronarias, por lo que actualmente se les está denominando "Compuestos Biológicamente Activos" (BAC según sus siglas en inglés) ya que, si bien carecen de valor nutritivo, no siempre resultan perjudiciales. Alguno de estos compuestos juegan un papel importante como defensa de la planta frente al ataque de todo tipo de depredadores y otros son compuestos de reserva que se acumulan en las semillas y serán utilizados a lo largo del proceso germinativo. Desde el punto de vista bioquímico los compuestos no-nutritivos son de naturaleza muy variada, no aparecen por igual en todas las plantas, sus efectos fisiológicos son distintos y por lo tanto su metodología de extracción, determinación y cuantificación tiene que ser también muy específica. Como ejemplo de algunos de estos Compuestos Biológicamente Activos de las leguminosas se pueden citar a aquellos de naturaleza proteica (inhibidores de proteasas, inhibidores de amilasas, lectinas) y los de naturaleza no proteica (glicósidos, aminoácidos libres, alcaloides, fitatos, fitoestrógenos, saponinas, taninos, etc.). Una de las vías para la disminución de estos compuestos cuando están en altas concentraciones es la mejora genética. Estos programas no solo se han centrado en conseguir una alta producción desde el punto de vista agronómico, sino de alcanzar una mejor calidad nutricional con la eliminación de estos componentes no deseables. Así, por ejemplo, existen variedades de distintas especies de Lupinus libres de alcaloides tóxicos. También se han obtenido variedades de Vicia faba sin vicina y convicina o con concentraciones muy bajas, así como Lathyrus sativus libres de un aminoácido neurotóxico.[45]
Se consumen por sus vainas tiernas o por sus granos: habas (Vicia faba), lentejas (Lens culinaris), garbanzos (Cicer arietinum), arvejas o guisantes (Pisum sativum), chícharo o muela (Lathyrus sativus); porotos, frijoles, habichuelas o judías (Phaseolus vulgaris, Ph. lunatus, Ph. coccineus y Ph. acutifolius), urd (Ph. mungo), frijol de vaca, caupí o cow-pea (Vigna sinensis), adzuki (V. angularis), poroto o frijol metro (Vigna sesquipedalis), soja o soya (Glycine max), poroto japonés (Dolichos lablab), guandú (Cajanus flavus), lupino o altramuz (Lupinus albus), tarhuí (Lupinus mutabilis), maní o cacahuete (Arachis hypogaea), guandsú o guisante de tierra (Voandzeia subterranea). Otras especies se utilizan en la alimentación por su fruto pulposo, como el pacay (Inga feuillei), el algarrobo blanco (Prosopis alba) el algarrobo europeo (Ceratonia siliqua) el algarrobo negro (Prosopis nigra) y el tamarindo (Tamarindus indica), entre otros. Aunque no es frecuente entre las leguminosas, hay especies cuya raíz carnosa o tubérculo se utiliza como alimento. Tal es el caso de la ajipa (Pachyrhizus ahipa), la jícama (Pachyrhizus erosus), Neocracca heterantha, Psoralea esculenta y Apios americana.
Finalmente, hay leguminosas que se utilizan para elaborar bebidas alcohólicas, tales como la "aloja de culén" que se prepara mediante infusión de tallos de culén (Psoralea glandulosa) con azúcar, granos de maíz tostado y varias especias. Los frutos de los algarrobos (Prosopis alba y P. nigra) se usan en el noroeste argentino para elaborar dos bebidas: la "añapa", bebida dulce y refrescante obtenida machacando los frutos en un mortero con agua, y la "aloja", bebida alcohólica autóctona obtenida por fermentación del mosto de las algarrobas que se hace moliendo los frutos con agua. Los frutos de ambos algarrobos también se usan para preparar dulces, tales como el "arrope" y el "patay".[46] El "café de Bonpland" o "café taperibá" se prepara con las semillas de Cassia occidentalis, torrándolas, y obteniendo una infusión similar al café, pero de sabor y aroma mucho más atenuados.[47]
Forrajeras
 Trébol frutilla en floración, asociado a gramíneas en una pradera.
Trébol frutilla en floración, asociado a gramíneas en una pradera.En la alimentación del ganado bovino y ovino, principalmente, las leguminosas -por sí solas o en asociación con las gramíneas forrajeras- presentan una serie de bondades que incrementan la producción de leche y carne y que, además, tienden a mejorar la eficiencia reproductiva de los rebaños. Algunas de tales características o ventajas son que constituyen una fuente importante de proteínas de buena calidad, dado que poseen una amplia gama de aminoácidos esenciales que las hacen superiores a las gramíneas. En este sentido, es de destacar que presentan una mayor concentración de nitrógeno en las hojas que las gramíneas. Presentan bajos niveles de fibras y alto contenido de calcio, en relación con las gramíneas. Por todo esto, las leguminosas forrajeras se siembran asociadas con gramíneas para mejorar la calidad de la oferta alimenticia para el ganado. No obstante, las leguminosas presentan un beneficio adicional como mejoradoras del suelo desde el punto de vista de la fertilidad, ya que tienen la propiedad de fijar el nitrógeno atmosférico en los nódulos radiculares. La fijación de nitrógeno que se realiza en estos nódulos, es aportado al suelo una vez han envejecido o muerto las raíces, siendo fácilmente aprovechado por otras plantas tales como las gramíneas con las que crecen asociadas. La cantidad de nitrógeno fijado por las leguminosas puede variar de 20 a 560 kg anuales por hectárea, dependiendo del tipo de suelo y de la humedad disponible. Esta particularidad de fijar nitrógeno le otorga a las leguminosas la facultad de habitar en suelos de fertilidad pobre, sin que esto afecte significativamente su producción y calidad de biomasa.[48]
Algunas de las especies de leguminosas utilizadas como forrajeras son la alfalfa (Medicago sativa), la lupulina (Medicago lupulina), los tréboles de carretilla (Medicago hispida, Medicago arabica y Medicago minima), los tréboles de olor (Melilotus alba, Melilotus officinalis), el trébol blanco ( Trifolium repens), el trébol rojo o de los prados ( Trifolium pratense), el trébol frutilla ( Trifolium fragiferum), el lotus o trébol pata de pájaro (Lotus tenuis, Lotus corniculatus), la mermelada de caballo (Desmodium discolor), la zulla (Hedysarum coronarium), las lespedezas (Lespedeza striata y Lespedeza stipulacea), las vicias o arvejillas (Vicia sativa, Vicia villosa), el caupí (Vigna sinensis), el kudzú (Pueraria javanica) y el gandul (Cajanus cajan) entre las más conocidas. Una de las principales limitaciones para el establecimiento y producción de las praderas de leguminosas anuales es la baja fertilidad de los suelos y, en particular, el bajo contenido de fósforo de los mismos. En ciertas zonas del oeste de Australia, por esta razón, se han introducido y desarrollado la serradela amarilla (Ornithopus compressus) y la serradela rosada (Ornithopus sativus) como plantas forrajeras, ya que se adaptan bien a suelos ácidos de texturas livianas, de baja fertilidad, y en especial pobres en fósforo.[49] [50] [51] Biserrula pelecinus es otra especie promisoria para el secano. Es una leguminosa endémica de la cuenca mediterránea de Europa y del norte de África. En su hábitat natural se adapta a suelos infértiles, de textura liviana, bien drenados y ligeramente ácidos.[52] El trébol balansa (Trifolium michelianum), originario de Turquía, es una especie que presenta una excelente adaptación a suelos con problemas de inundaciones.[53]
Melíferas
Las plantas melíferas son aquellas que ofrecen néctar a las abejas y avispas como recompensa por acarrear polen desde las flores de una planta a otra y, de ese modo, asegurar la polinización. Por extensión, también se incluyen todas las especies que estos insectos utilizan para recoger polen y propóleos. El factor de mayor importancia en cuanto a la atracción de las flores por las abejas es el néctar que secretan. La secreción alcanza su máximo durante el primer día de la apertura floral y va decreciendo paulatinamente. El néctar es una solución azucarada, con pequeñas cantidades de otras sustancias (aminoácidos, minerales, vitaminas, ácidos orgánicos, enzimas y aceites esenciales) que es secretado por órganos especializados de la planta denominados nectarios que generalmente suelen estar situados en la base de la corola —nectarios florales— y que se distinguen por su forma y color; en algunos casos están situados en las hojas o en los pedúnculos de las flores —nectarios extraflorales—. En general, las abejas manifiestan preferencia por el néctar que contiene más del 20% de azúcar.[54] Varias especies de leguminosas son buenas especies melíferas, tales como la alfalfa, el trébol blanco, el trébol de olor y varias especies de algarrobos.
Industriales
Gomas
Las gomas son exudados vegetales de carácter patológico, es decir, son el resultado de la agresión al vegetal por una picadura de insecto o una incisión natural o artificial. Esos exudados contienen polisacáridos heterogéneos, formados por diferentes azúcares y en general llevan ácidos urónicos. Se caracterizan por formar disoluciones coloidales viscosas. Hay diferentes especies productoras de gomas. Las más importantes están en la familia de las leguminosas. Se utilizan ampliamente en la industria farmacéutica, cosmética, alimentaria y textil. También son interesantes por sus propiedades terapéuticas; por ejemplo la goma arábiga es antitusígena y antiinflamatoria. Las más conocidas son la goma de tragacanto (Astragalus gummifer), la goma arábiga (Acacia senegal) y la goma guar (Cyamopsis tetragonoloba).[55]
Tintóreas
 Colorante índigo.
Colorante índigo.Entre las especies tintóreas se encuentran las siguientes. El palo de campeche Haematoxylon campechianum; árbol grande y espinoso que puede alcanzar hasta los 15 m de altura. La corteza es delgada y blanda y el corazón de la madera es sólido y duro. El corazón del tallo se utiliza para teñir y se logran colores rojos y morados. De esta especie se extrae el colorante histológico denominado hematoxilina. El palo de Brasil (Caesalpinia echinata) es un árbol muy parecido al anterior pero de menos altura y con flores rojizas o moradas. La madera se utiliza para teñir, también de rojo y morado. El guamuchil (Pithecallobium dulce) es otro árbol espinoso, que mide 4 m o más y presenta flores amarillentas o verdosas, dispuestas en cabezuelas. El fruto es rojizo y es el que se utiliza para teñir de color amarillo.[56] El colorante índigo se extrae del índigo verdadero Indigofera tinctoria, de Asia. En Centro y Sudamérica se extraen colorantes de dos especies afines a la anterior, el añil (Indigofera suffruticosa) y el índigo de Natal (Indigofera arrecta).
Ornamentales
 El ceibo es una de las tantas leguminosas utilizadas como plantas ornamentales. Además, es la Flor Nacional de Argentina y Uruguay.
El ceibo es una de las tantas leguminosas utilizadas como plantas ornamentales. Además, es la Flor Nacional de Argentina y Uruguay.Las leguminosas se han utilizado en todo el mundo como plantas ornamentales desde hace siglos. Una vasta diversidad de alturas, formas, colores de follaje, flores e inflorescencias hacen que esta familia sea muy utilizada en el diseño y parquización de jardines pequeños o de grandes parques.[12] A continuación se presenta una lista de las principales especies de leguminosas ornamentales, agrupadas por subfamilias:
- Subfamilia Caesalpinioideae: Bauhinia forficata, Caesalpinia gilliesii, Caesalpinia spinosa, Ceratonia siliqua, Cercis siliquastrum, Gleditsia triacanthos, Gymnocladus dioica, Parkinsonia aculeata, Senna multiglandulosa.[57]
- Subfamilia Mimosoideae: Acacia caven, Acacia cultriformis, Acacia dealbata, Acacia karroo, Acacia longifolia, Acacia melanoxylon, Acacia paradoxa, Acacia retinodes, Acacia saligna, Acacia verticillata, Acacia visco, Albizzia julibrissin, Calliandra tweediei, Paraserianthes lophantha, Prosopis chilensis.[57]
- Subfamilia Faboideae: Clianthus puniceus, Citysus scoparius, Erythrina crista-galli, Erythrina falcata, Laburnum anagyroides, Lotus peliorhynchus, Lupinus arboreus, Lupinus polyphyllus, Otholobium glandulosum, Retama monosperma, Robinia hispida, Robinia luxurians, Robinia pseudoacacia, Sophora japonica, Sophora macnabiana, Sophora macrocarpa, Spartium junceum, Teline monspessulana, Tipuana tipu, Wisteria sinensis.[57]
Leguminosas emblemáticas
- El ceibo (Erythrina crista-galli), es la flor nacional de Argentina y de Uruguay.[58]
- El guanacaste (Enterolobium cyclocarpum) es el árbol nacional de Costa Rica, por decreto del Poder Ejecutivo del 31 de agosto de 1959.[59]
- El palo brasil (Caesalpinia echinata) es el árbol nacional de Brasil desde 1978.[60]
- Acacia pycnantha es la flor nacional de Australia.[61]
- Bauhinia x blakeana es la flor nacional de Hong Kong.[62]
Referencias
- ↑ a b c d e f g Watson, L.; Dallwitz, M. J.. «Leguminosae» (en inglés). The families of flowering plants: descriptions, illustrations, identification, and information retrieval. Version: 1st June 2007.. Consultado el 9 de febrero de 2008.
- ↑ International Code of Botanical Nomenclature. En su artículo 18, inciso 5, expresa: "Los siguientes nombres, utilizados tradicionalmente, son tratados como válidamente publicados: ....Leguminosae (Fabaceae; tipo, Faba Mill. [= Vicia L.])....y establece que cuando las papilionáceas sean consideradas como una familia distinta a las restantes Leguminosas, el nombre Papilionaceae se conservará por sobre Leguminosae."
- ↑ a b c d e f g Judd, W. S., Campbell, C. S. Kellogg, E. A. Stevens, P.F. Donoghue, M. J. (2002), Plant systematics: a phylogenetic approach, Sinauer Axxoc, 287-292. ISBN 0-87893-403-0.
- ↑ a b c d e f g h i Stevens, P. F. (2001 en adelante). «Fabaceae» (en inglés). Angiosperm Phylogeny Website. Version 7 mayo 2006. Consultado el 28 de abril de 2008.
- ↑ Magallón, S. A., y Sanderson, M. J. (2001). «Absolute diversification rates in angiosperm clades» (en inglés). Evolution 55 (9): pp. 1762-1780. http://loco.biosci.arizona.edu/pdfs/MagallonEvolution2001.pdf.
- ↑ Burnham, R. J., & Johnson, K. R. 2004. South American palaeobotany and the origins of neotropical rain forests. Phil. Trans. Roy. Soc. London B, 359: 1595-1610.
- ↑ Lewis, G., B. Schrire, B. MacKinder, and M. Lock (eds). 2005. Legumes of the world. Royal Botanical Gardens, Kew, UK.
- ↑ Doyle, J. J., J. A. Chappill, C.D. Bailey, & T. Kajita. 2000. Towards a comprehensive phylogeny of legumes: evidence from rbcL sequences and non-molecular data. pp. 1 -20 in Advances in legume systematics, part 9, (P. S. Herendeen and A. Bruneau, eds.). Royal Botanic Gardens, Kew, UK.
- ↑ Kajita, T., H. Ohashi, Y. Tateishi, C. D. Bailey, and J. J. Doyle. 2001. rbcL and legume phylogeny, with particular reference to Phaseoleae, Millettieae, and allies. Systematic Botany 26: 515-536.
- ↑ Wojciechowski, M. F., M. Lavin y M. J. Sanderson (2004). «A phylogeny of legumes (Leguminosae) based on analysis of the plastid matK gene resolves many well-supported subclades within the family» (en inglés). American Journal of Botany 91: pp. 1846-1862. http://www.amjbot.org/cgi/content/full/91/11/1846.
- ↑ Angiosperm Phylogeny Group [APG] (2003). «An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG II» (en inglés). Botanical Journal of the Linnean Society 141: pp. 399-436. http://www.biodiversitas.org.br/floraBr/apg2.pdf.
- ↑ a b c d e f g h i j k l m Burkart, A. Leguminosas. En: Dimitri, M. 1987. Enciclopedia Argentina de Agricultura y Jardinería. Tomo I. Descripción de plantas cultivadas. Editorial ACME S.A.C.I., Buenos Aires. pag.: 467-538.
- ↑ a b Allen, O. N., & E. K. Allen. 1981. The Leguminosae, A Source Book of Characteristics, Uses, and Nodulation. The University of Wisconsin Press, Madison, USA.
- ↑ a b Lloret, L. & Martínez-Romero, E. 2005. Evolución y filogenia de Rhizobium. Vol. 47, No. 1-2 pp. 43 - 60. [1]
- ↑ a b c Sprent, J. I. 2001. Nodulation in legumes. Royal Botanic Gardens, Kew, UK.
- ↑ a b c Herendeen, P. S., W. L. Crepet, and D. L. Dilcher. 1992. The fossil history of the Leguminosae: phylogenetic and biogeographic implications. Pages 303 – 316 in Advances in Legume Systematics, part 4, the fossil record (P. S. Herendeen and D .L. Dilcher, eds). Royal Botanic Gardens, Kew, UK.
- ↑ Lewis, G., B. Schrire, B. MacKinder, and M. Lock (eds). 2005. Legumes of the world. Royal Botanical Gardens, Kew, UK.
- ↑ Crepet, W. L., and D. W. Taylor. 1985. The diversification of the Leguminosae: first fossil evidence of the Mimosoideae and Papilionoideae. Science 288: 1087-1089.
- ↑ Crepet, W. L., and D. W. Taylor. 1986. Primitive mimosoid flowers from the Paleocene-Eocene and their systematic and evolutionary implications. American J. Botany 73: 548-563.
- ↑ Crepet, W. L., and P. S. Herendeen. 1992. Papilionoid flowers from the early Eocene of southeastern North America. Pages 43–55 in Advances in Legume Systematics, part 4, the fossil record (P. S. Herendeen and D. L. Dilcher, eds.). Royal Botanic Gardens, Kew, UK.
- ↑ Herendeen, P. S. 1992. The fossil history of Leguminosae from the Eocene of southeastern North America. Pages 85-160 in Advances in Legume Systematics, part 4, the fossil record (Herendeen, P. S., and D. L. Dilcher, eds.). Royal Botanic Gardens, Kew, UK.
- ↑ Herendeen, P. S. 2001. The fossil record of the Leguminosae: recent advances. In Legumes Down Under: the Fourth International Legume conference, Abstracts, 34–35. Australian National University, Canberra, Australia.
- ↑ Herendeen, P. S., and S. Wing. 2001. Papilionoid legume fruits and leaves from the Paleocene of northwestern Wyoming. Botany 2001 Abstracts, published by Botanical Society of America (http://www.botany2001.org/).
- ↑ Wing, S. L., F. Herrera, and C. Jaramillo. 2004. A Paleocene flora from the Cerrajón Formation, Guajíra Peninsula, northeastern Colombia. Pages 146-147 in VII International Organization of Paleobotany Conference Abstracts (21-26 March). Museo Egidio Feruglio, Trelew, Argentina.
- ↑ Bruneau, A., Lewis, G. P., Herendeen, P. S., Schrire, B., & Mercure, M. 2008b. Biogeographic patterns in early-diverging clades of the Leguminosae. Pp. 98-99, in Botany 2008. Botany without Borders. [Botanical Society of America, Abstracts.]
- ↑ Bruneau, A., Mercure, M., Lewis, G. P., y Herendeen, P. S. (2008). «Phylogenetic patterns and diversification in the caesalpinioid legumes» (en inglés). Canadian Journal of Botany 86: pp. 697-718. doi:.
- ↑ Lavin, M., Herendeen, P. S., y Wojciechowski, M. F. (2005). «Evolutionary Rates Analysis of Leguminosae Implicates a Rapid Diversification of Lineages during the Tertiary» (en inglés). Systematic Biology 54 (4): pp. 575-594. doi:. http://sysbio.oxfordjournals.org/cgi/content/full/54/4/575.
- ↑ Wikström, N. et ál. (2001), Savolainen, V., & Chase, M. W. 2001. Evolution of the angiosperms: Calibrating the family tree. Proc. Roy. Soc. London B, 268: 2211-2220.
- ↑ Wojciechowski, M. F. 2003. Reconstructing the phylogeny of legumes (Leguminosae): An early 21st century perspective. Pp. 5-35, in Klitgaard, B. B. & Bruneau, A. (eds), Advances in Legume Systematics, Part 10, Higher Level Systematics. Royal Botanic Gardens, Kew.
- ↑ Wojciechowski, M. F. 2004. Astragalus (Fabaceae): A molecular phylogenetic perspective. Brittonia 57: 382-396.
- ↑ Wojciechowski, M. F. Sanderson, M. J., Baldwin, B. G., & Donoghue, M. J. 1993. Monophyly of aneuploid Astragalus: Evidence from nuclear ribosomal DNA internal transcribed spacer sequences. American J. Bot. 80: 711-722.
- ↑ a b Wojciechowski, Martin F., Johanna Mahn, and Bruce Jones. 2006. Fabaceae. legumes. Version 14 June 2006. The Tree of Life Web Project, http://tolweb.org/
- ↑ Käss, E., and M. Wink. 1996. Molecular evolution of the Leguminosae: phylogeny of the three subfamilies based on rbcL sequences. Biochemical Systematics and Evolution 24: 365-378.
- ↑ Käss, E., and M. Wink. 1997. Phylogenetic relationships in the Papilionoideae (Family Leguminosae) based on nucleotide sequences of cpDNA (rbcL) and ncDNA (ITS1 and 2). Molecular Phylogenetics and Evolution 8:65-88.
- ↑ Doyle, J.J., J.L. Doyle, J.A. Ballenger, E.E. Dickson, T. Kajita, and H. Ohashi. 1997. A phylogeny of the chloroplast gene rbcL in the Leguminosae: taxonomic correlations and insights into the evolution of nodulation. American J. Botany 84: 541-554.
- ↑ Lavin, M., J. J. Doyle, and J. D. Palmer. 1990. Evolutionary significance of the loss of the chloroplast--DNA inverted repeat in the Leguminosae subfamily Papilionoideae. Evolution 44: 390-402.
- ↑ Sanderson, M. J., and M. F. Wojciechowski. 1996. Diversification rates in a temperate legume clade: are there "so many species" of Astragalus (Fabaceae)? American J. Botany 83: 1488-1502.
- ↑ Chappill, J. A. 1995. Cladistic analysis of the Leguminosae: the development of an explicit hypothesis. Pages 1-10 in Advances in Legume Systematics, part 7, phylogeny (M. D. Crisp and J. J. Doyle, eds.). Royal Botanic Gardens, Kew, UK.
- ↑ Watson, L.; Dallwitz, M. J.. «The Genera of Leguminosae-Caesalpinioideae and Swartzieae» (en inglés). The families of flowering plants: descriptions, illustrations, identification, and information retrieval. Version: 1st June 2007.. Consultado el 11 de febrero de 2008.
- ↑ Gepts, P., W.D. Beavis, E.C. Brummer, R.C. Shoemaker, H.T. Stalker, N.F. Weeden, and N.D. Young. 2005. Legumes as a model plant family. Genomics for food and feed report of the cross-legume advances through genomics conference. Plant Physiol. 137: 1228 – 1235.
- ↑ Duke, J. A. 1992. Handbook of Legumes of Economic Importance. Plenum Press, New York, USA.
- ↑ Graham, P. H., & C. P. Vance. 2003. Legumes: importance and constraints to greater use. Plant Physiology 131: 872-877.
- ↑ Wojciechowski, M.F. 2006. Agriculturally & Economically Important Legumes.. Accedido el 15 de noviembre de 2008.
- ↑ «Information sheet: Pulses» (en inglés). The Vegetarian Society. Consultado el 9 de febrero de 2008.
- ↑ a b «Las leguminosas en la nutrición». Asociación Española de Leguminosas. Consultado el 9 de febrero de 2008.
- ↑ Carrizo, E. del V.; Palacio, M. O.; Roic, L.D.; Villaverde, A. A.; Soria Pereyra, M. L. y Torres, M.A. Utilización de especies de Prosopis L. por pobladores rurales en Santiago del Estero. Universidad Nacional de Santiago del Estero.
- ↑ Carrere, R.. «Productos alimenticios obtenibles de especies del monte indígena». El bosque natural uruguayo: utilización tradicional y usos alternativos. Grupo Guayubira. Consultado el 9 de febrero de 2008.
- ↑ Sánchez, A. (2005). «Leguminosas como potencial forrajero en la alimentación bovina». Revista de difusión de tecnología agrícola y pesquera del FONAIAP, Venezuela. Fonaiap Divulga, Versión Digital. Consultado el 17 de febrero de 2010.
- ↑ Paynter, B.H. 1990. Comparative phosphate requirements of yellow serradela (Ornithopus compressus), burr medic (Medicago polymorpha) and subterranean clover (Trifolium subterraneum). Aust. J. Exp. Agric. Anim. Husb. 30:507-514.
- ↑ Revell, C.K., and M. Ewing. 1994. Status of serradela (Ornithopus spp.) research in Western Australia. Alternative pasture legumes 1993. p. 47-49. Technical Report Nº219. In Michalk, D., Craig, A., and Collins, B. (eds.). Primary Industries South Australia, Australia.
- ↑ Revell, C.K., G.B. Taylor, and P.S. Cocks. 1999. Effect of length of growing season on development of hard seeds in yellow serradella and their subsequent softening at various depths of burial. Aust. J. Agric. Res. 50:1211-1223.
- ↑ Howieson, J.G., A. Loi, & S.J. Carr. 1995. Biserrula pelecinus L. a legume pasture species with potential for acid, duplex soils which is nodulated by unique root-nodule bacteria. Aust. J. Agric. Res. 46:997-1009.
- ↑ Dear, B., A. Craig y G. Sandral (2000). «Trifolium michelianum». NWS Agriculture Wagga Wagga Agricultural Institute Struan Research Center.
- ↑ Díaz González, T.E. Departamento de Biología de Organismos y Sistemas. Área de Botánica. Universidad de Oviedo. Plantas melíferas en Asturias. Importancia de las abejas para las plantas. [2]
- ↑ Kuklinski, C. 2000. Farmacognosia : estudio de las drogas y sustancias medicamentosas de origen natural. Ediciones Omega, Barcelona. ISBN 84-282-1191-4
- ↑ Marquez, A. C., Lara, O.F., Esquivel, R. B. & Mata, E. R. 1999. Composición, usos y actividad biológica: Plantas medicinales de México II. UNAM. Primera edición. México, D.F.
- ↑ a b c Macaya J. 1999. Leguminosas arbóreas y arbustivas cultivadas en Chile. Chloris Chilensis Año 2. Nº1.
- ↑ Ministerio de Educación de la Nación. Subsecretaría de Coordinación Administrativa. Día de la Flor Nacional "El Ceibo". Efemérides Culturales Argentinas. Consultado el 3 de marzo de 2010.
- ↑ Gilbert Vargas Ulate. 1997. Geografía turística de Costa Rica. EUNED, 180 p. ISBN 9977-64-900-6, 9789977649009.
- ↑ "Lei Nº 6.607, de 7 de dezembro de 1978. O Presidente da República, faço saber que o Congresso Nacional decreta e eu sanciono a seguinte Lei: Art. 1º- É declarada Árvore Nacional a leguminosa denominada Pau-Brasil (Caesalpinia echinata, Lam), cuja festa será comemorada, anualmente, quando o Ministério da Educação e Cultura promoverá campanha elucidativa sobre a relevância daquela espécie vegetal na História do Brasil."
- ↑ Boden, Anne (1985). «Golden Wattle: Floral Emblem of Australia» (http). Australian National Botanic Gardens.
- ↑ Williams, Martin (1999). «Golden Enigmatic Beauty» (http). Bahuninia.
Bibliografía
- Lewis G., Schrire B., Mackinder B. & Lock M. 2005. (eds.) Legumes of the world. The Royal Botanic Gardens, Kew, Reino Unido. 577 páginas. 2005. ISBN 1-900347-80-6.
Enlaces externos
 Wikiespecies tiene un artículo sobre Fabaceae. Wikispecies
Wikiespecies tiene un artículo sobre Fabaceae. Wikispecies Wikimedia Commons alberga contenido multimedia sobre Fabaceae. Commons
Wikimedia Commons alberga contenido multimedia sobre Fabaceae. Commons- Asociación Española de las Leguminosas (AEL). Entidad sin ánimo de lucro, constituida para la promoción de las leguminosas en España, entendiéndose como tales a los grupos botánicos que, perteneciendo a esta familia de plantas, sean de utilidad para la agricultura española.
- Faboideae Rudd (Papilionoideae Juss., es el nombre alternativo)











Wikimedia foundation. 2010.