- Haplogrupo B (ADNmt)
-
En genética mitocondrial humana el haplogrupo B uno de los grupos mitocondriales humanos (ADNmt). Se cree que apareció en el Extremo Oriente hace unos 50.000 años, se derivó del haplogrupo mitocondrial R y está determinado por los marcadores 8281-8289d y 16189.
En su libro The Seven Daughters of Eve, Bryan Sykes llamo a la mujer que originó este haplogrupo mitocondrial Ina.
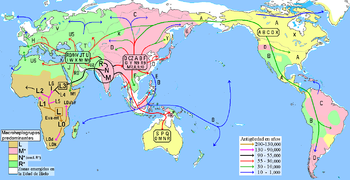 En poblaciones nativas de América y Extremo Oriente, zonas de predominio del Haplogrupo B en color verde.
En poblaciones nativas de América y Extremo Oriente, zonas de predominio del Haplogrupo B en color verde.
Contenido
Origen
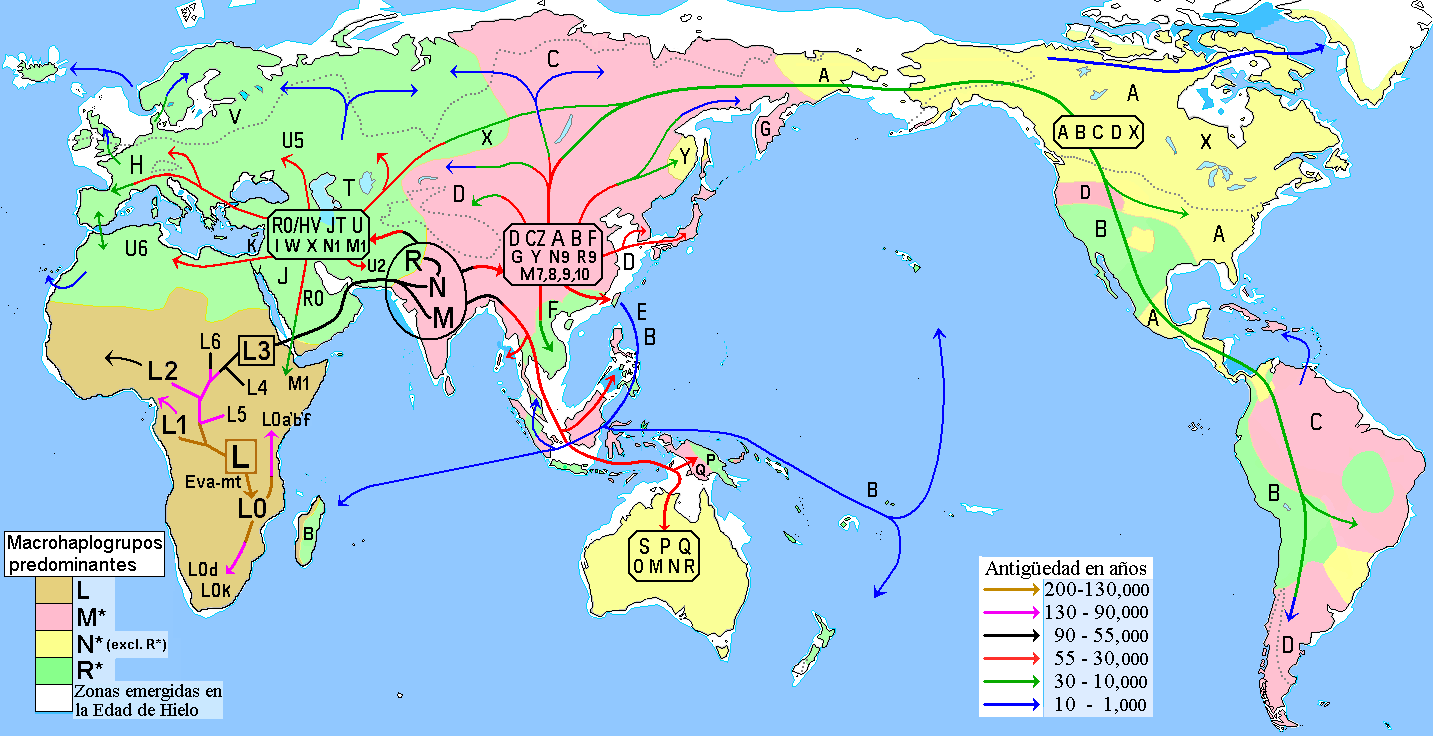
El haplogrupo B está relacionado con el haplogrupo R11, el cual está disperso en Oriente y parte de la India; así pues, B podría estar relacionado con los primeros pobladores que llegaron al Asia Oriental hace unos 50.000 años provenientes del Sur de Asia, atravesando zonas tropicales y de abundante vegetación en plena Edad de hielo. Tomando en cuenta que es en China donde se encuentra la mayor diversidad y población del haplogrupo B, éste se habría originado en el Extremo Oriente, probablemente entre el Sur de China y Sudeste de Asia.[1]
Otras versiones (véase enlaces externos), consideran a B originado en Asia Central hace 39.000–52.000 años. Pero esta teoría implica una migración en difíciles condiciones, pues para ello, los aún primitivos pobladores habrían tenido que recorrer una larga distancia, bordeando el Himalaya por el norte y atravesando zonas entre los desiertos árido y alpino, incluyendo los inviernos polares propios del clima y la geografía característica de la edad de hielo, por lo que esta vía no parece la más probable.
Distribución
Este haplogrupo es característico de todo Eurasia Oriental, América, Oceanía y Madagascar. El haplogrupo B se encuentra con mayor población en Asia Oriental.[2] El subgrupo B2 es uno de los 5 grupos haploides o haplogrupos que se encuentran en los pueblos indígenas de América. Los otros son los haplogrupos A, C, D y X.
Como los modelos de migración al Nuevo Mundo suponen que los primeros pobladores llegaron de Siberia, parecería sorprendente que el haplogrupo B fuera el único que no se encontrase en poblaciones siberianas actuales. Sin embargo, en realidad el haplogrupo B sí se ve entre siberianos del Sur, como los tuvanos, altayos, buriatos, mongoles, jakasios, telenguites y yakutos, aunque en bajas frecuencias;[3] y en algunos siberianos del norte como los evenkis.
Asia
- Asia Oriental: En China tiene un promedio aproximado del 17%, con las frecuencias más altas al Sur; destaca la ciudad de Cantón con 30%,[1] siendo también importante en Hong Kong (20%), Wuhan y Yunnan (19%). Fuera de China encontramos en aborígenes de Taiwán 32%,[4] han de Taiwán 17%, en Corea 14.5%[5] y Japón 13%.[6]
- Sudeste Asiático: Muy común en toda la región, tanto en aborígenes como en la población en general. Destaca las Filipinas con 33%[4] y es también común en Tailandia, Malasia, Indonesia, Indochina, en las islas indias de Nicobar, etc.
- Otras zonas de Asia: Muy extendido pero en bajas frecuencias en Asia Central y Sur de Siberia. Aisladamente en el Sur de Asia.
América
- Sudamérica: El haplogrupo B1 es típico de la zona andina quechua-aymara en una región equivalente a lo que fue el Imperio incaico. Las frecuencias más importantes están en el Altiplano andino, con un 88% en la región del Titicaca;[7] entre las poblaciones aymara se ha registrado una frecuencia de 74%.[8] El haplogrupo B se registra en comunidades aymara con frecuencias entre 67% y 93,9%[9] y en quechuas del Perú 37 a 68%.[10] En San Pedro de Atacama (Norte de Chile) 81%.[11] B es también predominante entre los wounaan[12] y otras poblaciones de la región colombiana del Pacifico[13] y en ciertas etnias en forma aislada, como en los yuco-yupka de Colombia,[12] oriundos de Churuguara (Venezuela) y xavantes del Mato Grosso (Brasil).
- Norteamérica: Típico en el llamado lejano oeste norteamericano, especialmente en el Arizona y Baja California en nativos kiliwa, mojave, yavapai y en general en hablantes de lenguas yumanas.[10]
Oceanía y Madagascar
Los navegantes austronesios colonizaron desde Indonesia las islas del Pacífico y Madagascar, expandiendo las lenguas malayo-polinesias y dispersando el haplogrupo B. Según promedio de algunas fuentes,[14] [15] las más altas frecuencias están en Polinesia con 98% y Micronesia 83%; encontrando en la isla de Nueva Guinea 20% y en las demás islas de Melanesia 36% como promedio.
Subclados
Haplogrupo B (8281-8289d)
- B4 (16217): Extendido en todo Extremo Oriente, Siberia, América y Oceanía.
- (16261)
- B4a: Importantes frecuencias al Sur de China (especialmente en Guangdong), Corea, Taiwán, Vietnam y Tailandia.
- B4a1
- B4a1a: En Extremo Oriente, especialmente en las Molucas (24,19%), algunas islas de Indonesia y en cuatro de las etnias aborígenes de Taiwán, alcanzando entre los amis del este de la isla una frecuencia de 44,9%.[16]
- B4a1b: En Corea
- B4a1c; En Asia Oriental y Sur de Siberia.
- B4a2: En Japón, Taiwán y Filipinas.
- B4a1
- B4g: Encontrado en China.
- B4a: Importantes frecuencias al Sur de China (especialmente en Guangdong), Corea, Taiwán, Vietnam y Tailandia.
- B4b'd'e (827, 15535)
- B4b: Asia Oriental y Sur de Siberia.
- B2: Exclusivo de los amerindios y extendido en Norte, Centro y Sudamérica. Alta frecuencia en la región andina sudamericana y zona Oeste de América del Norte.
- B4b1: Especialmente en aborígenes de Taiwán.
- B4d: En China y Siberia.
- B4e
- B4b: Asia Oriental y Sur de Siberia.
- B4c (3497, 16311)
- B4c1: Especialmente en Filipinas.
- B4c2: Encontrado en Uzbekistán, Tailandia y Tonga.
- B4f: En Ryukyu
- (16261)
- B5 (709, 8584, 9950, 10398, 16140): Extendido en todo Asia Oriental y Sudeste de Asia.
- B6 o B7: Encontrado en China.
Mutación paralela
Dentro del haplogrupo B se habría dado el caso de una mutación paralela coincidente, lo que dificulta su definición y filogenia, por lo que pudo haberse dado los siguientes casos:
- B estaría definido por la mutación 16189.[22] Esto significaría que la mutación 8281-8289d habría sido paralela según el siguiente esquema:[23]
B (16189) B4'5 (8281-8289d) B4 (16217)
B5
R11'B6 (12950) R11
B6 (8281-8289d)
R24
- B es comúnmente definido por la mutación 8281-8289d,[24] por lo que la mutación 12950 habría sido paralela del siguiente modo:
R11' B (16189) R11 (12950)
B (8281
-8289d)B4 (16217)
B5
B6 (12950)
R24
Véase también
Eva mitocondrial (L) L0 L1-6 L1 L2 L3 L4 L5 L6 M N CZ D E G Q A S R I W X Y C Z B F R0 JT P U HV J T K H V Enlaces externos
- Dispersión del Haplogrupo B, de la National Geographic
- Ina
- Haplogrupo B de Ian Logan
- Filogenia de R
Referencias
- ↑ a b Yong-Gang Yao et al. 2001, Phylogeographic Differentiation of Mitochondrial DNA in Han Chinese Am J Hum Genet. 2002 March; 70(3): 635–651
- ↑ Haplogroup B.
- ↑ M. Derenko et al. 2007, Phylogeographic Analysis of Mitochondrial DNA in Northern Asian Populations Am J Hum Genet. 2007 November; 81(5): 1025–1041
- ↑ a b Jean A. Trejaut et al. 2005, Traces of Archaic Mitochondrial Lineages Persist in Austronesian-Speaking Formosan Populations PLoS Biol 3(8): e247
- ↑ Han-Jun Jin et al. 2008,The Peopling of Korea Revealed by Analyses of Mitochondrial DNA and Y-Chromosomal Markers PLoS ONE. 2009; 4(1): e4210
- ↑ Tanaka M et al. 2004, Mitochondrial Genome Variation in Eastern Asia and the Peopling of Japan Genome Res. 2004. 14: 1832-1850
- ↑ José Sandoval et al. 2004, Variantes del ADNmt en isleños del lago Titicaca: máxima frecuencia del haplotipo B1 y evidencia de efecto fundador Rev. peru biol. v.11 n.2 Lima jul./dic. 2004
- ↑ Merriwether D.A. & R.E. Ferrell 1996. "The four founding lineage hypothesis: a critical re-evalulation". Molecular Phylogenetics and Evolution 51(1): 241-246.
- ↑ Lewis, Cecil M. Jr.; Raúl Y. Tito; Beatriz Lizárraga; and Anne C. Stone "Land, Language, and Loci: mtDNA in Native Americans and the Genetic History of Peru"; American Journal of Physical Anthropology 127:351–360.
- ↑ a b Citlalin Xochime 2002, People of the Red Brown Earth.
- ↑ a b Easton, Ruth D. et al 1996, mtDNA Variation in the Yanomami: Evidence for Additional New World Founding Lineages
- ↑ a b Keyeux, Genoveva; C. Rodas; N. Gélvez and D. Carter 2002. "Possible Migration Routes into South America Deduced from Mitochondrial DNA Studies in Colombian Amerindian Populations" Human Biology 74 (2): 211–233.
- ↑ Rondón, Francisco; J.C. Osorio; A.V. Peña; H.A. Garcés y G. Barreto "Diversidad genética en poblaciones humanas de dos regiones colombianas" Colombia Médica 39 (2) suplemento 2.
- ↑ D.A. Merriwether et al. 2005,Ancient mitochondrial M haplogroups identified in the Southwest Pacific. PNAS September 13, 2005 102 37 p13034-13039
- ↑ J. Friedlaender et al. 2005, Expanding Southwest Pacific mitochondrial haplogroups P and Q. MBE Advance Access published April 6, 2005
- ↑ Trejaut, Jean A. 2005. Traces of Archaic Mitochondrial Lineages Persist in Austronesian-Speaking Formosan Populations; PLoS Biology 3(8): e247.
- ↑ Friedlaender, Jonathan Scott 2007. Genes, language, and culture history in the Southwest Pacific: 72-75. Oxford University Press. ISBN 978-0-19-530030-7
- ↑ Kayser, Manfred et. al. 2006. Melanesian and Asian Origins of Polynesians: mtDNA and Y Chromosome Gradients Across the Pacific; Molecular Biology and Evolution 23(11): 2234-2244.
- ↑ Ricaut, François et al. 2009, A new deep branch of eurasian mtDNA macrohaplogroup M reveals additional complexity regarding the settlement of Madagascar BMC Genomics 2009, 10:605 doi:10.1186/1471-2164-10-605
- ↑ Mait Metspalu et al. 2004, Most of the extant mtDNA boundaries in South and Southwest Asia were likely shaped during the initial settlement of Eurasia by anatomically modern humans. BMC Genetics 2004, 5:26. doi:10.1186/1471-2156-5-26
- ↑ Catherine Hill et al. 2006, Phylogeography and Ethnogenesis of Aboriginal Southeast AsiansMol. Biol. Evol. 23(12):2480–2491. 2006
- ↑ Easton, Ruth D. et al 1996, mtDNA Variation in the Yanomami: Evidence for Additional New World Founding Lineages
- ↑ van Oven M, Kayser M. 2009, PhyloTree.org - mtDNA subtree R
- ↑ mtDNA Phylogenetic Tree Genebase.com
Categorías:- Haplogrupos mitocondriales humanos
- Genética humana
Wikimedia foundation. 2010.