- Micro ARN
-
Un micro ARN (miARN o miRNA por sus siglas en inglés) es un ARN monocatenario, de una longitud de entre 21 y 25 nucleótidos, y que tiene la capacidad de regular la expresión de otros genes mediante diversos procesos, utilizando para ello la ruta de ribointerferencia.[1]
Fueron descritos inicialmente en 1993 por Lee y colaboradores en el laboratorio de Victor Ambros,[2] sin embargo el término "microARN" sólo se acuñó en 2001 en un conjunto de tres artículos publicados en Science (26 Octubre 2001).[3] A principios de 2008, análisis computacionales realizados por IBM sugerían la presencia de alrededor de 50.000 miRNA diferentes en el genoma humano, cada uno tal vez con alrededor de miles de ARNm dianas potenciales.[4]
Contenido
Definición
Una vez descubiertos los siRNAs y la existencia en las células de proteínas que catalizan la degradación del ARNm, los investigadores se preguntaron si los siRNA también estaban codificados en el genoma, y empezaron a purificar pequeños ARNs (19-25 nt) a partir de diferentes especies animales. Sin embargo, no encontraron siRNA, sino los denominados micro RNA, que se habían identificado anteriormente de forma independiente.[2]
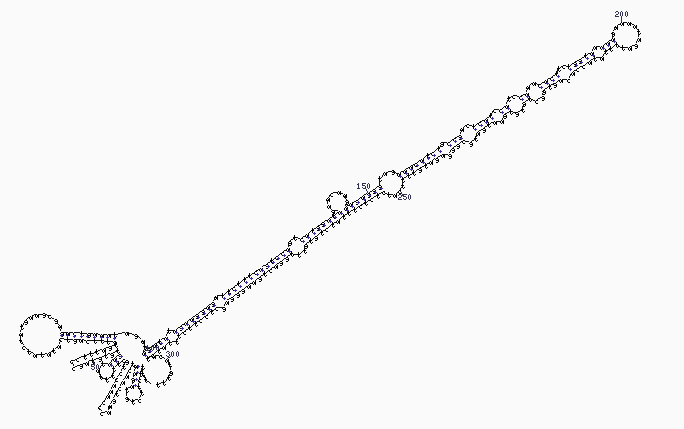
 La estructura secundaria en stem-loop (tallo-lazo) de un pre-miRNA de Brassica oleracea.
La estructura secundaria en stem-loop (tallo-lazo) de un pre-miRNA de Brassica oleracea.
Los miRNA son moléculas de ARN transcritas a partir de genes de ADN, pero no son traducidas a proteínas. Se expresan en una amplia variedad de organismos, desde plantas hasta gusanos y humanos. Muchos miRNA están bien conservados entre especies,[6] y muchos componentes de la maquinaria de los miRNA se han encontrado incluso en Archaea y eubacterias, lo que revela que su origen es muy antiguo. Algunos recuentos de miRNA en humanos identificaban hasta 800, lo que implicaría que los miRNA podrían representar como mínimo el 3% de todos los genes humanos.[7]
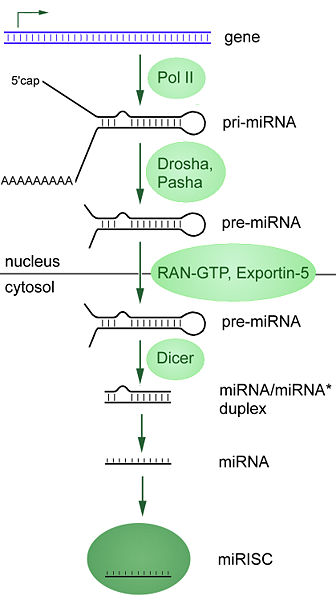
La secuencia de ADN que codifica para un gen de miRNA tiene una longitud que supera al tamaño final del propio miRNA e incluye la región miRNA y una región que es complementaria a la anterior, lo que permite su apareamiento. Esto conlleva que, durante la transcripción de esta secuencia de ADN, se forman regiones que tienen la capacidad de formar una horquilla y generar un ARN bicatenario primario largo conocido como pri-miRNA. Posteriormente, un enzima nuclear llamado drosha corta las bases de la horquilla, formando lo que se denomina pre-miRNA. Este pre-miRNA es transportado desde el núcleo al citoplasma por la exportina 5. Una vez que el pre-miRNA está en el citoplasma es fragmentado por la enzima dicer, que lo corta hasta la longitud final de 20-25 nucleotidos.[5]
Función
La función de los miRNA está relacionada con la regulación de la expresión génica. De esta forma un miRNA es complementario de una parte de uno o más ARN mensajeros (ARNm). Los miRNA de animales suelen mostrar complementariedad imperfecta con la región 3' UTR, y generalmente inhiben la traducción del ARNm, mientras que los de plantas suelen mostrar complementariedad perfecta con regiones codificantes e inducen el corte y la posterior degradación del ARNm diana (como ocurre con los siRNAs en animales).[5]
Antes de clasificarlos como parte de la ruta del RNAi, los miRNA fueron identificados inicialmente en gusanos, en los que regulan las fases del desarrollo,[8] pero en la actualidad se sabe que están implicados en una amplia variedad de procesos del desarrollo y podrían tener una función en el establecimiento de redes y en el ajuste fino de la expresión génica en la célula.[9] [6] Dado que el número de dianas potenciales de los miRNA aumenta al número de miles (alrededor del 30% de los genes humanos), los miRNA podrían constituir otra capa del circuito regulatorio que existe en las células.[10] Según esto, cualquier desregulación de los miRNA podría conllevar grandes problemas regulatorios en la célula, induciendo quizá fenotipos cancerosos. De hecho, se ha mostrado que los perfiles de expresión de los miRNA están modificados en un gran número de tipos de cáncer[11] y que la sobreexpresión forzada de los miRNA podría conducir al desarrollo de tumores.[12]
Características de los miRNA
Los miRNA se transcriben a partir de diferentes localizaciones genómicas como largos tránscritos primarios (pri-miRNA) por la ARN-polimerasa II.[13]
Los miRNA pueden encontrarse en muchos tipos de loci en el genoma:[14]
- La transcripción de los miRNA puede estar regulada independientemente por promotores específicos, como ocurre con miR-1–1 y miR-133a-2, que están regulados por los factores de transcripción SRF (Serum Response Factor) y MyoD[15] (ver el apartado "Funciones durante el desarrollo" para más detalles). Tales miRNA pueden de todas maneras estar localizados en intrones pero a menudo con una orientación antisense.
- La mayoría de los precursores de miRNA presentes en intrones tienen la misma orientación que el gen en el que se alojan y son inicialmente transcritos como parte de su ARN precursor.[6] Por ejemplo, miR-208, que se expresa específicamente en el corazón de humano y ratón, reside en el intrón 28 de la cadena pesada de la α-miosina cardiaca. Los pri-miRNA intrónicos pueden producirse por corte del intrón por la endonucleasa Drosha, después de que se haya producido el proceso de splicing.
- Con pocas excepciones, los miRNA que están integrados o se superponen con exones de tránscritos conocidos, están siempre en la misma orientación, y la mayoría se encuentran en las regiones no-codificantes UTRs 5′ o 3′ (por ejemplo, miR-198 en follistatin-like 1).
Aproximadamente el 50% de los miRNA están en clusters de miRNA que están inicialmente codificados como un tránscrito policistrónico (que incluye varios genes),[16] que posteriormente se fragmenta en múltiples miRNA. En la mayoría de los casos, los miRNA policistrónicos comparten el mismo patrón de expresión. Sin embargo, los niveles relativos de los miRNA dentro del cluster parecen estar regulados de una manera dependiente del desarrollo y la homeostasis, lo que sugiere una complejidad aún no definida en la regulación de la expresión génica.
Biogénesis y procesamiento
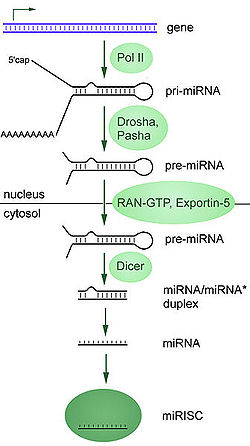 Los microRNA (miRNA) se producen a partir de un microARN precursor (pre-miRNA), que a su vez se forma a partir de un tránscrito de microARN primario (pri-miRNA).
Los microRNA (miRNA) se producen a partir de un microARN precursor (pre-miRNA), que a su vez se forma a partir de un tránscrito de microARN primario (pri-miRNA).Los genes que codifican miRNA son mucho más largos que los miRNA procesados maduros; los miRNA se transcriben inicialmente como tránscritos primarios o pri-miRNA con una caperuza en 5' y una cola de poli-adeninas (poly-A) en 3' y se procesan en el núcleo celular en estructuras cortas de 70-nucleótidos en forma de tallo-lazo (stem-loop) conocidas como pre-miRNA. En animales este procesamiento se realiza por un complejo proteico llamado Microprocesador, que consta de la nucleasa Drosha[17] y la proteína de unión a ARN de doble hélice Pasha.[18] [19] Estos pre-miRNA son luego procesados a miRNA maduros en el citoplasma mediante la interacción con la endonucleasa Dicer, que también inicia la formación del complejo RISC (RNA-induced silencing complex).[20] Este complejo es el responsable del silenciamiento de genes que se observa debido a la expresión de los miRNA y a la interferencia de ARN mediada por siRNAs. La ruta varía ligeramente en plantas, debido a que carecen de homólogos de Drosha; en su lugar, sólo homólogos de Dicer realizan algunos de los pasos del procesamiento.[21] La ruta también es diferente para los miRNA derivados de tallos-lazos (stem-loops) procedentes de secuencias intrónicas; en este caso se procesan por Dicer pero no por Drosha.[22] Tanto la hebra sense como la antisense del ADN pueden funcionar como molde para producir miRNA.[23]
Drosha[24] (RNASEN en humanos)[25] es una proteína nuclear de un tamaño entre 130 y 160 kDa (kiloDalton). Contiene los dominios siguientes:
- dos dominios ARNasa III (los dominios catalíticos)
- un dominio dsRBD (double-strand RNA-binding domain, de unión al ARN doble hebra)
- dominios conservados (en la parte N-terminal), de los que no se conoce la función.
Drosha funciona en un complejo (Microprocesador), conjuntamente con una proteína de unión a ARN (denominada Pasha en Drosophila o DGCR8 en mamíferos). DGCR8[26] [27] es una proteína de unión a ADN que reconoce la zona de unión dsRNA–ssRNA (ARN doble hebra-simple hebra) y posiciona a la ribonucleasa Drosha a una distancia de 11 nucleótidos (que corresponde a una vuelta de hélice) desde la zona de unión. Por tanto, la función de DGCR8 en el complejo Microprocesador es análoga al dominio PAZ de Dicer; DGCR8 proporciona especificidad de sustrato y posiciona adecuadamente el centro de la ribonucleasa Drosha. Sin embargo, en el caso de Drosha-DGCR8, el dominio de especificidad está localizado en una cadena polipeptídica separada de los dominios ARNasa III. Se ha propuesto que el dominio WW de unión a prolinas de DGCR8 interacciona con el extremo N-terminal de Drosha, rico en prolinas. Es posible que Drosha tenga otras posibles proteínas asociadas que presenten dominios WW que aporten especificidades de sustrato alternativas y funciones biológicas adicionales de Drosha.
El procesamiento eficiente de los pre-miRNA por Drosha requiere la presencia de largas colas de ARN de hebra simple tanto en el extremo 3' como 5' de la molécula en horquilla.[28] Estos motivos de ARN de hebra simple (ssRNA) pueden variar en composicion, mientras que su longitud es de gran importancia para que el procesamiento tenga lugar. Un análisis bioinformático de pri-miRNA en humanos y moscas identificó regiones estructurales similares, denominadas 'segmentos basales (basal segments)', 'tallos inferiores (lower stems)', 'tallos superiores (upper stems)' y 'lazos terminales (terminal loops)'; basándose en estas estructuras conservadas, se han determinado perfiles termodinámicos de los pri-miRNA.[29] El complejo de Drosha (Microprocesador) corta la molécula de ARN ~2 vueltas de hélice a partir del lazo terminal y ~1 vuelta de hélice a partir de los segmentos basales. En la mayoría de las moléculas analizadas esta región contiene nucleótidos no apareados y la energía libre del duplex es relativamente alta en comparación con las regiones tallo superior e inferior [cita requerida]. La mayor parte de los pre-miRNA no presentan una estructura de ARN doble hebra (dsRNA) perfecta con un lazo terminal final. Existen algunas explicaciones posibles para esta selectividad. Una podría ser que dsRNA más largos de 21 pares de bases activan la respuesta de interferón y la maquinaria antiviral de la célula. Otra explicación plausible podría ser que el perfil termodinámico del pre-miRNA determina qué hebra será incorporada en el complejo Dicer. De hecho, se han detectado claras similitudes entre pri-miRNA codificados bien en hebras 5' o 3'.[29]
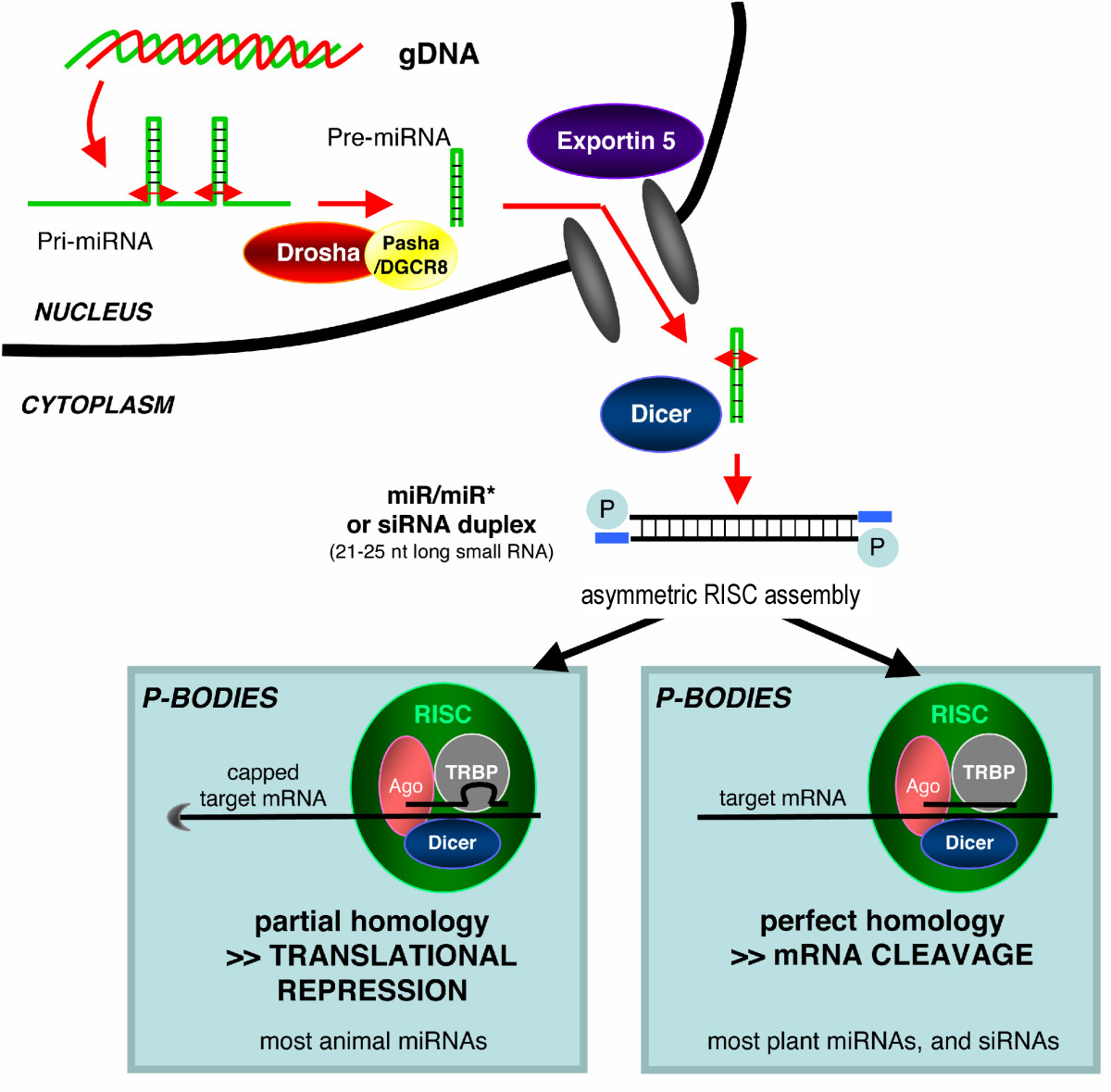
Una vez generado el pre-miRNA, Dicer corta el tallo-lazo (stem-loop) y se forman dos moléculas complementarias cortas, pero sólo una se integra en el complejo RISC (la antisense), como ocurre con los siRNAs. Esta hebra se conoce como la hebra guía, que es seleccionada por la proteína Argonauta, la ARNasa catalíticamente activa en el complejo RISC, en función de la estabilidad del extremo 5'.[30] La otra hebra (la sense), conocida como anti-guía o hebra pasajera, es degradada por el complejo RISC.[31] Después de su integración en el complejo RISC, ahora activado (ver las notas sobre el complejo RISC en siRNAs), los miRNA se emparejan de acuerdo con su secuencia de bases con la molécula de ARNm complementaria, y en animales, a diferencia con los siRNA, en la mayoría de los casos inducen la inhibición de la traducción de dicho ARNm.
Sin embargo, a pesar de que Dicer es una enzima fundamental en el procesamiento de los microRNA, se ha identificado una ruta de biogénesis de miRNA independiente de Dicer que utiliza la actividad catalítica de corte de Argonauta2 (Ago2). Así, a diferencia de otros miRNA, los niveles de algunos miRNA (miR-451-5', miR-2190-5', miR-2190-3', y miR-735-5') no se alteran en mutantes con pérdida de función de Dicer, pero disminuyen en mutantes MZago2 (maternal-zygotic). En el caso de miR-451 (un miRNA implicado en la diferenciación de la línea eritroide[32] ), el procesamiento de pre-miR-451 requiere la actividad catalítica de Ago2 in vivo. Los mutantes MZago2 muestran un retraso en la eritropoyesis que puede rescatarse utilizando Ago2 de tipo salvaje o con duplex de miR-451, pero no con Ago2 catalíticamente inactiva. Por ello, se ha sugerido que Ago2 es capaz de generar miRNA funcionales independientemente de Dicer.[33] Resultados similares se han observado en organismos diferentes.[34]
Como se indica en el caso de los siRNA, todavía no está claro cómo el complejo RISC activado localiza los ARNm complementarios en el interior celular. Las proteínas Argonauta, los componentes catalíticos de RISC, están localizadas en regiones específicas del citoplasma denominadas P-bodies (cuerpos-P, también cuerpos citoplásmicos o cuerpos GW, porque contienen la proteína GW182), los cuales son regiones con altas tasas de degradación de ARNm;[35] también se ha detectado actividad miRNA en los P-bodies.[36] Alteración de los P-bodies disminuye la eficiencia del proceso de RNAi, lo que sugería que los P-bodies podrían ser un lugar crítico para el proceso de RNAi.[37] Sin embargo, estudios posteriores han demostrado que los P-bodies no son imprescindibles para el proceso de RNAi, ya que células sin P-bodies pueden producir silenciamiento tanto con siRNA como con miRNA.[38]
Modo de funcionamiento
A pesar del importante progreso realizado en la comprensión de la biogénesis y la función de los miRNA, los mecanismos utilizados por los miRNA para regular la expresión génica permanecen bajo un intenso debate.[39] En efecto, existen trabajos publicados que indican que los miRNA en células animales reprimen la expresión génica de cuatro formas diferentes:
- degradación de la proteína durante la traducción
- inhibición de la elongación de la traducción
- terminación prematura de la traducción (disgregación de los ribosomas)
- inhibición de la iniciación de la traducción
Además, los miRNA en animales pueden inducir una degradación significativa de los ARNm diana (como los miRNA de plantas), a pesar del apareamiento imperfecto ARNm-miRNA. Sin embargo, el mecanismo de degradación suele ser diferente: los miRNA inducen la degradación de los ARNm diana mediante la eliminación de la caperuza (en el extremo 5') y de la cola de poliadeninas (poly-A, en el extremo 3').[40] Finalmente, los microARNs podrían también silenciar sus ARNm dianas secuestrándolos en foci (sitios) citoplásmicos discretos, los cuerpos de procesamiento de ARNm o P bodies, que carecen de maquinaria de traducción. Sin embargo, a pesar de las discrepancias existentes entre los diferentes mecanismos propuestos, los apoyos experimentales para cada mecanismo son variados, y son el objeto actual de intensos estudios, para tratar de elucidarlas. Se ha sugerido que las diferencias observadas se deben a deficiencias en los experimentos realizados, en algunos casos originadas por la utilización de modelos erróneos en los estudios de regulación de la traducción.[41]
Por último, en estudios recientes se ha detectado que en determinadas condiciones, los miRNA pueden también activar la síntesis proteica.[42]
Características generales de los miRNA:[39]
- se asocian a la región 3’ UTR de los ARNm diana.
- hacen falta muchos sitios de unión para activar la respuesta de los miRNA (la unión de uno solo no produce efectos significativos).
- un ARNm puede estar regulado por diferentes miRNA.
- un único miRNA puede controlar la actividad de cientos de ARNm diferentes.[43] Los primeros estudios a gran escala publicados en Nature en 2008, que utilizan una variante de la técnica de espectrometría de masas denominada SILAC (marcado estable de aminoácidos con isótopos pesados en cultivo celular, Stable Isotope Labelling with Amino acids in Cell culture) para detectar las proteínas afectadas al reducir o aumentar los niveles de un miRNA concreto, muestran que en efecto, un único miRNA puede reducir los niveles de cientos de proteínas mediante el bloqueo de su traducción, no sólo degradando sus ARNm.[44] [45] El descubrimiento más llamativo de estos estudios es que los efectos de los miRNA sobre las proteínas son habitualmente bastante modestos, modificando sus niveles de expresión tan sólo en un factor 2. Sin embargo, a pesar de que los efectos de los miRNA pueden ser sutiles, también pueden ser potentes: los miRNA parecen intervenir en la regulación de la expresión génica realizando un ajuste fino, de forma compleja e interconectada.
- la especificidad y la función de los miRNA están determinados por los nucleótidos 2 a 7 de la parte 5' de los miRNA maduros (la llamada región "semilla" del miRNA): dichos nts deben ser obligatoriamente complementarios al ARNm diana.[46]
- un miRNA puede ser funcional aunque no haya sido sintetizado en el núcleo: un miRNA introducido en la célula por transfección puede inhibir eficazmente la síntesis de proteínas. En este caso, como ocurre con los siRNAs, el efecto del miRNA se puede modular extracelularmente utilizando moléculas químicas pequeñas. En un estudio publicado en Agosto 2008, se desarrolló un ensayo basado en células para monitorizar la actividad de la ruta del RNAi, y los autores detectaron que la molécula enoxacin (Penetrex) mejora la degradación del ARNm mediada por siRNA, y promueve la biogénesis de los miRNA endógenos.[47] Este artículo prueba que pequeñas moléculas químicas pueden mejorar la eficacia de la ruta del RNAi, lo que podría ser útil en el desarrollo de herramientas para la investigación y terapéuticas.
- el miRNA es el responsable de la especificiad de sustrato (el ARNm diana) del miRNP (complejo micro-ribo-nucleo-proteico, el miRNA unido al complejo RISC).
Función biológica
Función en la infección viral y la respuesta inmune
- Función anti-viral: el miRNA miR-32 tiene como diana una secuencia presente en la UTR del extremo 3’ de todos los ARNm retrovirales[48]
- Función pro-viral: el miRNA miR-122, que se expresa específicamente en el hígado, es necesario para que el virus de la hepatitis C se exprese de manera eficiente.[49]
- Muchos miRNA que están regulados diferencialmente en líneas celulares hematopoyéticas tienen funciones importantes en la regulación del desarrollo y la función de las células del sistema inmune, y en las interacciones huésped-patógeno.[50]
Función en cáncer
Los miRNA pueden funcionar como supresores de tumores o como oncogenes; queda por demostrar su influencia concreta en cada tipo de cáncer.[51] De hecho, un estudio mostró que cerca del 50% de los miRNA anotados en humanos están localizados en áreas del genoma conocidas como sitios frágiles,[52] que están asociadas con el desarrollo de cáncer.
La expresión de ciertos miRNA está correlacionada con varios tipos de cáncer, por lo que funcionarían como oncogenes. Por ejemplo, un informe de Sonoki y colaboradores[53] relacionó el gen mir-125b-1 con leucemia, y describió un paciente con leucemia linfoblástica aguda de precursores de células B que portaba una inserción del pre-miRNA en el locus de la cadena pesada de la inmunoglobulina. Aunque los investigadores no pudieron determinar cómo se modulaba la expresión de mir-125b-1 en las células tumorales, este estudio apoya el papel de este gen como un oncomir.
La primera indicación de que los miRNA podrían funcionar como supresores de tumores proceden de un informe de Calin y colaboradores[54] que mostraba que pacientes diagnosticados con una forma frecuente de leucemia en adultos (leucemia linfoide crónica de las células B o LLC), presentan a menudo deleciones o sub-regulación (downregulation) de dos genes de miRNA presentes en un cluster, mir-15a y mir-16-1. Deleciones dentro del locus 13q14 ocurren en más del 65% de los casos de LLC, y en más del 50% de los linfomas de las células del manto, el 16–40% de los mielomas múltiples y el 60% de los cánceres de próstata. Por tanto, se predijo que un gen supresor de tumores debía residir en esta región de 30-kb. Es interesante notar que mir-15a y mir-16-1 mapean dentro del intrón de un gen de ARN no-codificante para proteína, de función desconocida, denominado LEU2. Por otro lado, algunos estudios establecen una conexión entre la reducción de la expresión de let-7 (que regula la proliferación y diferenciación celular en C. elegans) y el incremento de la tumorigénesis y el pronóstico grave de los pacientes afectados.[55] Además, durante el desarrollo normal, LIN28 (un promotor de pluripotencia) puede prevenir la acumulación de let-7. De acuerdo con estos resultados, se ha propuesto que let-7 regularía la capacidad pluripotente (stemness) de las células, reprimiendo la auto-renovación y promoviendo la diferenciación, tanto durante el desarrollo normal como en cáncer.
Por otro lado, se ha observado además que los microRNA contribuyen a la progresión maligna del cáncer, específicamente mediando la invasión tumoral y la formación de metástasis.[56]
Función durante el desarrollo
Por ejemplo, en C. elegans, los miRNA permiten un paso rápido a través de las diferentes fases del desarrollo:[57] los miRNA lin-4[58] y let-7[59] controlan el momento en los que se define el destino de las células neuronales e hipodérmicas durante el desarrollo larvario.[60] lin-4, let-7 y otros genes de miRNA están consevados en mamíferos, y su función potencial en el desarrollo embrionario de mamíferos están bajo estudio activo. En C. elegans, los niveles de expresión de LIN-14[61] y LIN-28[62] disminuyen debido a la expresión del ARN de lin-4 al final del primer estadio larvario, para permitir la progresión a estadios laravarios posteriores. En los estadios larvarios finales, la expresión de LIN-41[63] y otros genes podría verse reducida por la expresión del ARN de let-7, liberando la represión de la expresión de la proteína LIN-29[64] y permitiendo la progresión al estadio adulto. Dado que el ARNm de lin-29 no posee sitios complementarios al ARN de let-7, probablemente lin-29 no es una diana directa de let-7.
Por otro lado, en miocitos de mamíferos, la activación del factor de transcripción SRF[65] induce la expresión de miR-1–1[66] y miR-1–2,[67] que a su vez reprime la expresión del factor de transcripción Hand2[68] y el ligando de Notch,[69] Delta, afectando por tanto la expansión de las células progenitoras o su diferenciación.[70] SRF también induce la expresión de miR-133a-1[71] y miR-133a-2,[72] lo que inhibe a su vez SRF en un lazo de retroalimentación (feedback loop). En músculo opera una ruta similar, excepto que la inhibición por retroalimentación de Mef2[73] y MyoD[74] ocurre cuando la expresión de HDAC4[75] está reducida debido al silenciamiento ejercido por miR-1–1 y miR-1–2.
Uno de los primeros miRNA detectados con una función en el desarrollo del sistema inmune[50] fue miR-181a;[76] este miRNA se expresa en altos niveles en las células del timo y en niveles menores en las células del corazón, los nódulos linfáticos y la médula ósea. En la médula ósea, miR-181a se expresa en niveles mayores por células B B220+ B que por células T CD3+. Específicamente, la expresión de miR-181a en células B derivadas de la médula ósea disminuye durante la maduración de las células B desde el estadio del desarrollo de pro-célula-B al estadio pre-célula-B. Además, la expresión ectópica de miR-181a en células enriquecidas en células madre y progenitoras hematopoyéticas resultó en un incremento en el porcentaje de células B CD19+ y un descenso en el porcentaje de células T CD8+ en ensayos de reconstitución de la médula ósea a corto plazo en ratones, demostrando que la especificidad de líneas celulares de los miRNA podría tener una función en la regulación del desarrollo de los linfocitos.
Por otro lado, también se ha detectado una distribución espacial de los miRNA: en embriones del pez cebra (zebrafish), los patrones de localización de miRNA individuales indican que su actividad podría estar limitada a los tejidos y órganos en los que se expresan. Por ejemplo, miR-206[77] se expresa sobre todo en el músculo, miR-126[78] en los vasos sanguíneos y el corazón, miR-200a[79] en el sistema de la línea lateral (un sistema mecanosensorial que detecta el movimiento del agua) y órganos sensoriales, y miR-30c[80] en el precursor de los riñones.[81]
Nuevas funciones maternas
Dos estudios independientes en 2007 en ratones[82] [83] indican que una cantidad significativa de miRNA maternos se heredan por los zigotos, y que dichos miRNA maternos podrían tener un papel importante en las etapas tempranas del desarrollo embrionario (efecto materno).
Referencias
- ↑ Pillai RS. MicroRNA function: multiple mechanisms for a tiny RNA? RNA 2005 Dec;11(12):1753-6 [1]
- ↑ a b Lee RC, Feinbaum RL, Ambros V (1993). «The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14.». Cell 75: pp. 843–854. doi:. PMID 8252621.
- ↑ Ruvkun, G. (Oct 26 2001). «Molecular biology. Glimpses of a tiny RNA world.». Science 294 (5543): pp. 797–9. doi:. PMID 11679654. http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=pubmed&dopt=Abstract&list_uids=11679654.
- ↑ Glasser V. Tapping miRNA-Regulated Pathways. Genetic Eng Biotech News Mar 1 2008 (Vol. 28, No. 5) [2]
- ↑ a b c Saumet A, Lecellier CH (2006). «Anti-viral RNA silencing: do we look like plants?». Retrovirology 3 (3): pp. 3. doi:. PMID 16409629.
- ↑ a b c Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 2004 Jan 23;116(2):281-97 [3]
- ↑ Bentwich I, Avniel A, Karov Y, Aharonov R, Gilad S, Barad O, Barzilai A, Einat P, Einav U, Meiri E, Sharon E, Spector Y, Bentwich Z. Identification of hundreds of conserved and nonconserved human microRNAs. Nat Genet. 2005 Jul;37(7):766-70 [4]
- ↑ Wightman B, Ha I, Ruvkun G. Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell 1993 Dec 3;75(5):855-62 [5]
- ↑ Ambros V. MicroRNA pathways in flies and worms: growth, death, fat, stress, and timing. Cell 2003 Jun 13;113(6):673-6[6]
- ↑ Bartel DP, Chen CZ. Micromanagers of gene expression: the potentially widespread influence of metazoan microRNA. Nat Rev Genet. 2004 May;5(5):396-400[7]
- ↑ Lu C, Tej SS, Luo S, Haudenschild CD, Meyers BC, Green PJ. Elucidation of the small RNA component of the transcriptome. Science 2005 Sep 2;309(5740):1567-9[8]
- ↑ He L, Thomson JM, Hemann MT, Hernando-Monge E, Mu D, Goodson S, Powers S, Cordon-Cardo C, Lowe SW, Hannon GJ, Hammond SM. A microRNA polycistron as a potential human oncogene. Nature 2005 Jun 9;435(7043):828-33[9]
- ↑ Kim VN. MicroRNA biogenesis: coordinated cropping and dicing. Nat Rev Mol Cell Biol. 2005 May;6(5):376-85[10]
- ↑ Zhao Y, Srivastava D.A developmental view of microRNA function. Trends Biochem Sci. 2007 Apr;32(4):189-97[11]
- ↑ Zhao Y, Samal E, Srivastava D. Serum response factor regulates a muscle-specific microRNA that targets Hand2 during cardiogenesis. Nature 2005 Jul 14;436(7048):214-20 [12]
- ↑ O'Donnell KA, Wentzel EA, Zeller KI, Dang CV, Mendell JT. c-Myc-regulated microRNAs modulate E2F1 expression. Nature 2005 Jun 9;435(7043):839-43 [13]
- ↑ Drosha [14]
- ↑ Pasha [15]
- ↑ Denli AM, Tops BB, Plasterk RH, Ketting RF, Hannon GJ. (2004). Nature 432(7014):231-5.
- ↑ Bernstein E, Caudy AA, Hammond SM, Hannon GJ. (2001). Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature 409(6818):363-6.
- ↑ Kurihara Y, Watanabe Y. (2004). Arabidopsis micro-RNA biogenesis through Dicer-like 1 protein functions. Proc Natl Acad Sci USA 101(34):12753-8.
- ↑ Gao F-B (2007). «Posttranscriptional control of neuronal development by microRNA networks». Trends in Neurosciences 31: pp. 20. doi:.
- ↑ Stark A, Bushati N, Jan CH, et al (2008). «A single Hox locus in Drosophila produces functional microRNA from opposite DNA strands». Genes Dev. 22 (1): pp. 8–13. doi:. PMID 18172160.
- ↑ Filipowicz W, Jaskiewicz L, Kolb FA, Pillai RS. Post-transcriptional gene silencing by siRNAs and miRNAs. Curr Opin Struct Biol. 2005 Jun;15(3):331-41 [16]
- ↑ RNASEN [17]
- ↑ MacRae IJ, Doudna JA. Ribonuclease revisited: structural insights into ribonuclease III family enzymes. Curr Opin Struct Biol. 2007 Feb;17(1):138-45 [18]
- ↑ DGCR8 [19]
- ↑ Zeng Y, Cullen BR (2005). «Efficient processing of primary microRNA hairpins by Drosha requires flanking nonstructured RNA sequences». J. Biol. Chem. 280 (30): pp. 27595–603. doi:. PMID 15932881. http://www.jbc.org/cgi/content/full/280/30/27595.
- ↑ a b Han J, Lee Y, Yeom K-H, Nam J-W, Heo I, Rhee J-K, Sohn SY, Cho Y, Zhang B-T, Kim VN (2006). «Molecular basis for the recognition of primary microRNAs by the Drosha-DGCR8 complex». Cell 125 (5): pp. 887–901. doi:. http://www.cell.com/content/article/abstract?uid=PIIS0092867406005162.
- ↑ Preall JB, He Z, Gorra JM, Sontheimer EJ. (2006). Short interfering RNA strand selection is independent of dsRNA processing polarity during RNAi in Drosophila. Curr Biol 16(5):530-5.
- ↑ Gregory RI, Chendrimada TP, Cooch N, Shiekhattar R. (2005). Human RISC couples microRNA biogenesis and posttranscriptional gene silencing. Cell 123(4):631-40.
- ↑ Zhan, M. and Miller, C.P. and Papayannopoulou, T. and Stamatoyannopoulos, G. and Song, C.Z.}, (2007). «MicroRNA expression dynamics during murine and human erythroid differentiation». Experimental hematology 35 (7): pp. 1015-1025. doi:. PMID 17588470.
- ↑ Cifuentes, D. and Xue, H. and Taylor, D.W. and Patnode, H. and Mishima, Y. and Cheloufi, S. and Ma, E. and Mane, S. and Hannon, G.J. and Lawson, N. and others (2010). «A novel miRNA processing pathway independent of dicer requires Argonaute2 catalytic activity». Science 328 (5986): pp. 1694 - 1698. doi:.
- ↑ Heng-Chi Lee, Liande Li, Weifeng Gu, Zhihong Xue, Susan K. Crosthwaite, Alexander Pertsemlidis, Zachary A. Lewis, Michael Freitag, Eric U. Selker, Craig C. Mello, Yi Liu (2010). «Diverse Pathways Generate MicroRNA-like RNAs and Dicer-Independent Small Interfering RNAs in Fungi». Molecular Cell 38 (6): pp. 803-814. doi:.
- ↑ Sen G, Blau H (2005). «Argonaute 2/RISC resides in sites of mammalian mRNA decay known as cytoplasmic bodies». Nat Cell Biol 7 (6): pp. 633–6. doi:. PMID 15908945.
- ↑ Lian S, Jakymiw A, Eystathioy T, Hamel J, Fritzler M, Chan E (2006). «GW bodies, microRNAs and the cell cycle». Cell Cycle 5 (3): pp. 242–5. PMID 16418578.
- ↑ Jakymiw A, Lian S, Eystathioy T, Li S, Satoh M, Hamel J, Fritzler M, Chan E (2005). «Disruption of P bodies impairs mammalian RNA interference». Nat Cell Biol 7 (12): pp. 1267–74. doi:. PMID 16284622.
- ↑ Eulalio A, Behm-Ansmant I, Schweizer D, Izaurralde E. P-body formation is a consequence, not the cause, of RNA-mediated gene silencing. Mol Cell Biol. 2007 Jun;27(11):3970-81 [20]
- ↑ a b Eulalio A, Huntzinger E, Izaurralde E. Getting to the root of miRNA-mediated gene silencing. Cell 2008 Jan 11;132(1):9-14 [21]
- ↑ Eulalio A, Rehwinkel J, Stricker M, Huntzinger E, Yang SF, Doerks T, Dorner S, Bork P, Boutros M, Izaurralde E. Target-specific requirements for enhancers of decapping in miRNA-mediated gene silencing. Genes Dev. 2007 Oct 15;21(20):2558-70[22]
- ↑ Kozak M. (2008). «Faulty old ideas about translational regulation paved the way for current confusion about how microRNAs function.». Gene Jul 22 (Epub ahead of print). [23]
- ↑ Vasudevan S, Tong Y, Steitz JA. Switching from repression to activation: microRNAs can up-regulate translation. Science 2007 Dec 21;318(5858):1931-4 [24]
- ↑ Hayden EC (2008). «Thousands of proteins affected by miRNA». Nature 545. 562. [25]
- ↑ Selbach M, Schwanhäusser B, Thierfelder N, Fang Z, Khanin R, Rajewsky N. (2008). «Widespread changes in protein synthesis induced by microRNAs». Nature Published online 30 July 2008. [26]
- ↑ Baek D, Villén J, Shin C, Camargo FD, Gygi SP, Bartel DP. (2008). «The impact of microRNAs on protein output». Nature Published online 30 July 2008. [27]
- ↑ Lewis BP, Burge CB, Bartel DP. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell 2005 Jan 14;120(1):15-20 [28]
- ↑ Shan G, Li Y, Zhang J, Li W, Szulwach KE, Duan R, Faghihi MA, Khalil AM, Lu L, Paroo Z, Chan AW, Shi Z, Liu Q, Wahlestedt C, He C, Jin P. (2008). «A small molecule enhances RNA interference and promotes microRNA processing.». Nat Biotechnol. 26 (8). 933-40. [29]
- ↑ Lecellier CH, Dunoyer P, Arar K, Lehmann-Che J, Eyquem S, Himber C, Saïb A, Voinnet O. A cellular microRNA mediates antiviral defense in human cells. Science 2005 Apr 22;308(5721):557-60 [30]
- ↑ ="Jopling2005" Jopling CL, Yi M, Lancaster AM, Lemon SM, Sarnow P. Modulation of hepatitis C virus RNA abundance by a liver-specific MicroRNA. Science 2005 Sep 2;309(5740):1577-81 [31]
- ↑ a b Lodish HF, Zhou B, Liu G, Chen CZ. Micromanagement of the immune system by microRNAs. Nat Rev Immunol. 2008 Feb;8(2):120-30 [32]
- ↑ Esquela-Kerscher A, Slack FJ. Oncomirs - microRNAs with a role in cancer. Nat Rev Cancer. 2006 Apr;6(4):259-69 [33]
- ↑ Calin GA, Sevignani C, Dumitru CD, Hyslop T, Noch E, Yendamuri S, Shimizu M, Rattan S, Bullrich F, Negrini M, Croce CM. Human microRNA genes are frequently located at fragile sites and genomic regions involved in cancers. Proc Natl Acad Sci U S A 2004 Mar 2;101(9):2999-3004 [34]
- ↑ Sonoki T, Iwanaga E, Mitsuya H, Asou N.Insertion of microRNA-125b-1, a human homologue of lin-4, into a rearranged immunoglobulin heavy chain gene locus in a patient with precursor B-cell acute lymphoblastic leukemia. Leukemia 2005 Nov;19(11):2009-10 [35]
- ↑ Calin GA, Dumitru CD, Shimizu M, Bichi R, Zupo S, Noch E, Aldler H, Rattan S, Keating M, Rai K, Rassenti L, Kipps T, Negrini M, Bullrich F, Croce CM. Frequent deletions and down-regulation of micro- RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia. Proc Natl Acad Sci U S A. 2002 Nov 26;99(24):15524-9 [36]
- ↑ Büssing I, Slack FJ, Großhans H. (2008). «let-7 microRNA in development, stem cells and cancer». Trends Mol Med. Jul 30. (Epub ahead of print). [37]
- ↑ Ma L, Weinberg RA. (2008). «Micromanagers of malignancy: role of microRNAs in regulating metastasis». Trends Genet. Jul 30 (Epub ahead of print). [38]
- ↑ Lau NC, Lim LP, Weinstein EG, Bartel DP. An abundant class of tiny RNAs with probable regulatory roles in Caenorhabditis elegans. Science 2001 Oct 26;294(5543):858-62 [39]
- ↑ lin-4 [40]
- ↑ let-7 [41]
- ↑ Reinhart BJ, Slack FJ, Basson M, Pasquinelli AE, Bettinger JC, Rougvie AE, Horvitz HR, Ruvkun G. The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature 2000 Feb 24;403(6772):901-6 [42]
- ↑ LIN-14 [43]
- ↑ LIN28 [44]
- ↑ LIN-41 [45]
- ↑ LIN-29 [46]
- ↑ SRF [47]
- ↑ miR-1-1 [48]
- ↑ miR-1-2 [49]
- ↑ Hand2 [50]
- ↑ Notch [51]
- ↑ Zhao Y, Srivastava D. A developmental view of microRNA function. Trends Biochem Sci. 2007 Apr;32(4):189-97 [52]
- ↑ miR-133a-1 [53]
- ↑ miR-133a-2 [54]
- ↑ Mef2 [55]
- ↑ MyoD [56]
- ↑ HDAC4 [57]
- ↑ miR-181-a [58]
- ↑ miR-206 [59]
- ↑ miR-126 [60]
- ↑ miR-200a [61]
- ↑ miR-30c [62]
- ↑ Wienholds E, Kloosterman WP, Miska E, Alvarez-Saavedra E, Berezikov E, de Bruijn E, Horvitz HR, Kauppinen S, Plasterk RH. MicroRNA expression in zebrafish embryonic development. Science 2005 Jul 8;309(5732):310-1 [63]
- ↑ Murchison EP, Stein P, Xuan Z, Pan H, Zhang MQ, Schultz RM, Hannon GJ. Critical roles for Dicer in the female germline. Genes Dev. 2007 Mar 15;21(6):682-93 [64]
- ↑ Tang F, Kaneda M, O'Carroll D, Hajkova P, Barton SC, Sun YA, Lee C, Tarakhovsky A, Lao K, Surani MA. Maternal microRNAs are essential for mouse zygotic development. Genes Dev. 2007 Mar 15;21(6):644-8 [65]
Categoría:- ARN
Wikimedia foundation. 2010.