- Punto de control
-
Punto de control
Los puntos de control (checkpoints en inglés) son mecanismos moleculares (no necesariamente agregados moleculares) que verifican que se cumplen las condiciones necesarias para permitir el paso de una fase del ciclo celular a otra, impidiendo así que ciertos eventos como daños en el ácido desoxirribonucleico (ADN) trasciendan a lo largo del ciclo.
Contenido
Puntos de control en el ciclo celular
Los puntos de control celular son mecanismos que aseguran la fidelidad de la división celular en las células eucariotas. Tales puntos de control verifican si los procesos en cada fase del ciclo celular han sido completados con precisión antes de progresar hacia la siguiente fase. Han sido identificados múltiples puntos de control, aunque algunos son mejor conocidos que otros.
En 1986, Temple y Raff describieron el ciclo celular como un reloj;[1] si éste fuera el caso, cada una de las fases funcionaría de acuerdo a una especie de reloj interno, que determinaría cuánto tiempo debería durar. Sin embargo, actualmente el ciclo celular se describe como las piezas de un dominó que, al caer, hacen que la siguiente caiga también: igualmente, para que una fase del ciclo celular tenga lugar, la fase anterior tiene que haber finalizado correctamente. Los puntos de control aseguran que una fase haya finalizado antes de pasar a la siguiente. Por tanto, los checkpoints son mecanismos de control que refuerzan la dependencia durante el ciclo celular.[2] Los sucesos del ciclo celular de la mayor parte de los organismos están organizados en rutas dependientes, en los cuales el inicio de los sucesos tardíos depende de que los sucesos iniciales hayan finalizado correctamente. La existencia de un mecanismo de control se evidencia cuando una droga, un mutante u otra condición libera una relación de dependencia en el ciclo celular: el suceso secundario en una ruta determinada tiene lugar aunque no se hayan cumplido los prerrequisitos necesarios. Por ejemplo, para que tenga lugar la mitosis es necesario que se haya completado la replicación del ADN, pero esta dependencia puede eliminarse mediante mutación de proteínas concretas, de forma que la mitosis puede ocurrir aunque el ADN no haya terminado de replicarse. Esto implica que la dependencia se debe a la existencia de un mecanismo de control (un checkpoint en el ciclo celular) y no a una característica intrínseca de los propios procesos.
Una función importante de muchos puntos de control consiste en evaluar los daños en el ADN, los cuales se detectan por mecanismos sensores. Cuando se localiza el daño, el punto de control envía una señal que detiene el ciclo celular hasta que se realiza la reparación o, cuando no es posible repararlo, marca la célula para su destrucción por apoptosis (mecanismo efector). Todos los checkpoints que valoran daños en el ADN parece que utilizan el mismo mecanismo sensor-señal-efector.
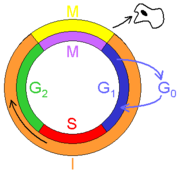 Ciclo celular. M: mitosis, I: interfase.
Ciclo celular. M: mitosis, I: interfase.
La mayor parte de las células de un organismo adulto están diferenciadas y no se dividen. Algunos tipos celulares están diferenciados de forma terminal y no pueden volver a proliferar a lo largo de toda la vida del individuo: es el caso de las células del músculo esquelético y cardíaco, los adipocitos y las neuronas. Otros tipos celulares, sin embargo, permanecen en estado quiescente (G0) pero pueden ser estimuladas para re-entrar en el ciclo celular, como los linfocitos T y B, y los fibroblastos. Para ello, antes de entrar en fase S deben superar un punto de control denominado "punto de restricción" que se encuentra al final de la fase G1. Esto se consigue mediante el incremento en los niveles de ciclinas D inducido por los factores de crecimiento. Por el contrario, las células que tienen capacidad de división (las células madre) deben cesar de dividirse, salir del ciclo celular y entrar en la fase fase G0 para comenzar el proceso de diferenciación y convertirse en células con una función especializada. Las únicas células que se dividen continuamente en el humano adulto son las células madre hematopoyéticas y las células epiteliales del intestino.
Los principales puntos de control que verifican la progresión a través del ciclo celular en eucariotas son los siguientes:
Punto de Restricción
Es el primer checkpoint del ciclo celular, al final de la fase G1, justo antes de entrar en la fase S.[3] La mayor parte de las células se paran en este momento y entran en un estado de reposo denominado G0. Las células eucarióticas normalmente se detienen en este punto de control si las condiciones ambientales son adversas (falta de nutrientes, por ejemplo). En células animales este punto de control se denomina "punto de restricción", mientras que en levaduras se denomina "punto de inicio" (start).
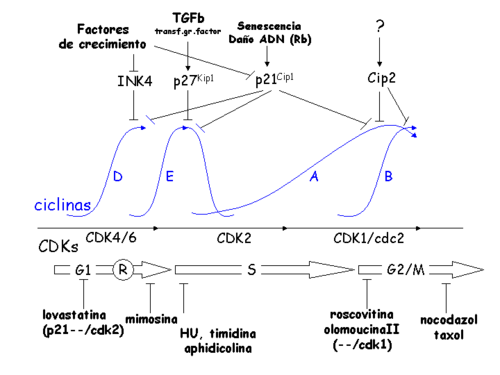 Esquema que muestra la variación en los niveles de ciclinas y CDKs a lo largo del ciclo celular, así como la acción de diversos inhibidores.
Esquema que muestra la variación en los niveles de ciclinas y CDKs a lo largo del ciclo celular, así como la acción de diversos inhibidores.Este momento en G1 fue descrito por primera vez en 1974 por Arthur Pardee, quien lo denominó "punto de restricción" R.[4] Pardee observó que las células que han pasado el punto R pueden progresar a través de la fase S independientemente de la presencia de mitógenos.[5] Además, Pardee identificó que este checkpoint no funcionaba correctamente en líneas celulares cancerosas. Las líneas celulares cancerosas que se utilizaron en este estudio estaban infectadas con el virus de simio 40 (SV40).[4] El descubrimiento de que las proteínas oncogénicas de los virus tumorales (como el antígeno grande T de SV40, E1A de adenovirus y E7 de HPV), desactivan el checkpoint de G1/S mediante su interacción inhibidora con el producto del gen de retinoblastoma,[6] [7] proporcionó datos fundamentales para la comprensión de los mecanismos moleculares que subyacen en este mecanismo de control.
Este checkpoint está controlado fundamentalmente por la acción de los inhibidores de CDKs (CKI) denominados p16 (también INK4A) y p21Waf1.[8] Estas proteínas inhiben CDK4/6 y aseguran que no pueda interaccionar con la ciclina D1 para continuar el avance en el ciclo celular. En condiciones de crecimiento inducido o expresión oncogénica de ciclina D, este checkpoint se supera porque la expresión aumentada de ciclina D permite su interacción con CDK4/6. Una vez que se forman los complejos activos CDK4/6-ciclina D, éstos fosforilan la proteína supresora de tumores retinoblastoma (Rb) y otras proteínas de la misma familia (p130 y p107), lo que libera la inhibición del factor de transcripción denominado E2F. Esto a su vez genera la expresión de moléculas requeridas para la transición G1/S, como las ciclinas E y A, la timidina kinasa, la timidilato sintetasa, la ADN polimerasa α o la dihidrofolato reductasa (DHFR), entre otras. Las ciclinas E y A interaccionan y activan la quinasa dependiente de ciclina 2 (CDK2), el principal activador de la transición G1/S, que impulsa el paso hacia la fase S.
Checkpoint de daños en el ADN
La variación de la secuencia del ADN en la línea germinal es esencial para mantener la variabilidad genética y asegurar la aparición de modificaciones que permitan una mejor adaptación al medio. Sin embargo, en la línea somática los cambios genéticos normalmente son nocivos, y las células han desarrollado estrictos mecanismos de seguridad para detectar y corregir las posibles alteraciones que haya podido sufrir el ADN. Una única alteración genómica (una mutación) que produzca una simple modificación en la cantidad producida de una proteína, o la sustitución de un único aminoácido en su secuencia, puede desencadenar una serie de variaciones que culminen con la generación de un tumor. La aparición de este tipo de errores están asociados con los fallos en los mecanismos que deberían haber detectado y corregido la lesión en el ADN que causó la mutación, o en último término, dirigido la célula afectada hacia un proceso de muerte celular. Por ello, los mecanismos que aseguran la integridad del ADN son fundamentales para el correcto funcionamiento celular.
Las modificaciones en la secuencia del ADN pueden generarse por modificaciones químicas espontáneas de sus componentes, por errores durante la replicación o por daños infligidos al ADN, debido a la presencia de agentes endógenos (generados por el propio metabolismo celular normal) o exógenos (procedentes del exterior). Entre éstos se encuentran la radiación ionizante (IR) y determinados productos químicos (véase Daño del ADN). El daño en el ADN inicia una respuesta que activa diferentes mecanismos de reparación que reconocen lesiones específicas en el ADN, que son reparadas en el momento para recuperar la secuencia original del ADN. Asimismo, el daño en el ADN provoca una parada en el ciclo celular, que conlleva la alteración de numerosos procesos fisiológicos, que a su vez implica síntesis, transporte y degradación de proteínas. La comprensión de cuál es la señal que emana de las lesiones en el ADN y cómo afecta a las numerosas rutas celulares implicadas se ha obtenido en gran parte estudiando la sofisticada respuesta a las roturas de doble hebra (DSBs, por Double Strand Breaks), que está conservada desde levaduras hasta humanos. Esta respuesta es una intricada red de señalización movilizada fundamentalmente por dos proteínas kinasas asociadas: ATM (Ataxia-telangiectasia Mutated, el gen mutado en esta enfermedad humana) y ATR (ATM- and Rad3- Related).[9] [10]
Transición G1/S
Este checkpoint impide la preparación para la replicación del ADN hasta que se haya eliminado el daño detectado en el ADN. Uno de los eventos clave en este proceso es la activación inducida por el daño en el ADN de la molécula p53. La activación de p53 incluye su fosforilación, acetilación y sumolación, además de su estabilización y su translocación hacia el núcleo celular. La fosforilación de p53 perturba su interacción con la ligasa E3 de ubiquitina hMdm2, lo que impide la ubiquitinación y degradación de p53. La principal responsable de la fosforilación de p53 en diferentes aminoácidos es ATM, que además contribuye a la estabilización de p53 mediante la fosforilación de hMdm2 (que inhibe su interacción con p53) y del factor de transcripción E2F1 (que activa la trascripción de p19ARF, un inhibidor de hMdm2). Una vez estabilizada, p53 entra en el núcleo celular e induce la transcripción de varios genes, entre los cuales se incluyen el inhibidor de kinasas dependientes de ciclinas (CDKs) denominado p21Waf1. Esta molécula inhibe la formación de los complejos ciclina D-CDK4, lo que impide la transición G1/S. Simultáneamente, p53 induce la transcripción de genes implicados en la reparación del ADN.
Alternativamente, si el daño genómico es demasiado grande para que pueda ser reparado, ATM induce la activación de una serie de rutas celulares que culminan en la muerte celular.
Intra S
ATM juega también un papel importante en el control de la respuesta al daño al ADN inducido por IR durante la fase S. En este caso, ATM fosforila e induce hChk2, que a su vez fosforila Cdc25A. Esta es una proteína que defosforila tirosinas y que, en células no irradiadas, defosforila CDK2 para promover la transición de G1 a S. Cuando hChk2 está activado y fosforila Cdc25A, promueve la degradación de esta fosfatasa por el proteasoma, lo que impide la activación de los complejos ciclina E-CDK2, provocando así una parada en la iniciación y progresión de la replicación del ADN.
Además, la activación de ATM inducida por IR produce la fosforilación de la proteína BRCA1, lo cual produce la disociación de BRCA1 de los complejos en los que se encuentra. BRCA1 se asocia a muchas proteínas que están implicadas en la respuesta al daño al ADN y hay múltiples evidencias que sugieren que BRCA1 realiza funciones múltiples en este proceso a través de sus proteínas asociadas. Entre éstas, se encuentran proteínas implicadas en modificaciones de la cromatina (SWI/SNF, HDAC1/2, p300/CBP), proteínas implicadas en la reparación del ADN (BACH1, BLM...) y proteínas implicadas en el control de la transcripción (GADD45, BARD1...).[11] BRCA1 es el gen de susceptibilidad al cáncer de mama y ovario, mutado en el 50% de todos los casos familiares de estos tipos de cáncer.
Transición G2/M
El checkpoint de G2/M evita que entren en mitosis células que o bien han terminado la replicación de su ADN y han sido posteriormente expuestas a agentes que lo han dañado, o bien células que han traspasado el checkpoint intra-S con daño en el ADN sin reparar.
ATM también es importante para el funcionamiento de este checkpoint. Como en el caso anterior, ATM es necesario para la fosforilación de hChk2, y hChk2 es necesario para el mantenimiento del checkpoint de G2/M. hChk2 fosforila e inhibe la fosfatasa hCdc25C, que por tanto no puede defosforilar y activar los complejos ciclina B1-CDK1, que son necesarios para promover la entrada en mitosis.
Por otro lado, aunque ATM es la primera molécula en activarse en la respuesta de daño al ADN, y es la responsable de la respuesta rápida e inmediata, ATR se une en una fase tardía y mantiene el estado fosforilado de sustratos específicos. Esta importante redundancia añade una mayor complejidad a la respuesta. Sin embargo, además de su papel en la fase tardía de la respuesta, ATR también responde a daños al ADN que no activan ATM, como tratamientos de radiación UV, horquillas de replicación bloqueadas e hipoxia, defosforilando al menos algunos de los sustratos de ATM, como p53 y BRCA1.[9] [10]
Checkpoint del ensamblaje del huso
Durante la mitosis, la célula organiza un huso mitótico con dos polos, al cual se anclan los cromosomas mediante unas estructuras proteicas denominadas cinetocoros. Los cromosomas constan de dos moléculas idénticas de ADN, las cromátidas hermanas, que permanecen unidas hasta anafase a través de unas proteínas denominadas cohesinas. El mecanismo que detecta que se ha formado correctamente un huso mitótico, que todos los cromosomas están asociados a dicho huso de manera bipolar, y que todos ellos se encuentran alineados en la placa metafásica es el denominado checkpoint de mitosis, de anafase o también punto de control del ensamblaje del huso, abreviado SAC por sus siglas en inglés (Spindle Assembly Checkpoint).[12]
Si uno de los cromosomas, por alguna razón, se retrasa durante el proceso de alineamiento, esta maquinaria produce una detención temporal de la progresión en el ciclo celular : la célula se detiene en metafase, dando tiempo a los mecanismos de reparación a resolver el problema detectado. Si pasado un tiempo, el problema no se ha corregido, la célula será abocada a un proceso de muerte celular, un mecanismo de seguridad para evitar que se produzca una situación de aneuploidía, generalmente con consecuencias graves para el organismo.
Cuando todos los cromosomas se congregan correctamente en la placa metafásica, el checkpoint de mitosis se inactiva, de manera que se produce el corte de las cohesinas que mantenían unidas las cromátidas hermanas, disparándose de este modo la entrada en anafase. Finalmente, ambos juegos de cromátidas hermanas se organizan alrededor de los polos de las células hijas en telofase, y por tanto ambas células reciben un complemento cromosómico completo.
Referencias
- ↑ Temple, S.; Raff, M.C. (1986), "Clonal analysis of oligodendrocyte development in culture: evidence for a developmental clock that …", Cell(Cambridge) 44 (5): 773–779 [1]
- ↑ Hartwell L.H., Weinert T.A. (1989). «Checkpoints: controls that ensure the order of cell cycle events.» Science. Vol. 246. n.º 4930. 629-634. [2]
- ↑ Foijer, F.; Te Riele, H. (2006), "Restriction beyond the restriction point: mitogen requirement for G2 passage", Cell Division 1 (1): 8, doi:, http://celldiv.com/content/1/1/8
- ↑ a b Pardee, A.B. (1974), "A Restriction Point for Control of Normal Animal Cell Proliferation", Proceedings of the National Academy of Sciences 71 (4): 1286–1290, doi:, http://www.pnas.org/cgi/reprint/71/4/1286.pdf
- ↑ Rossow, P.W.; Riddle, V.G.H.; Pardee, A.B. (1979), "Synthesis of Labile, Serum-Dependent Protein in Early G1 Controls Animal Cell Growth", Proceedings of the National Academy of Sciences 76 (9): 4446–4450, doi:, http://www.pnas.org/cgi/reprint/76/9/4446.pdf
- ↑ Pardee, A.B. (1989), "G1 events and regulation of cell proliferation", Science 246 (4930): 603–608, doi:, http://www.sciencemag.org/cgi/content/abstract/246/4930/603
- ↑ Munger, K.; Phelps, W.C.; Bubb, V.; Howley, P.M.; Schlegel, R. (1989), "The E6 and E7 genes of the human papillomavirus type 16 together are necessary and sufficient for …", Journal of Virology 63 (10): 4417–4421, http://jvi.asm.org/cgi/content/abstract/63/10/4417
- ↑ Malumbres, Marcos; Barbacid, Mariano (2001), "[http://www.sbs.utexas.edu/genetics/Literature/articles/NatureCellDivisionMilestones/ToCycleOrNot-Cancer.pdf Milestones in cell division To cycle or not to cycle: a critical decision in cancer]", Nature Reviews Cancer 1 (3): 222–231, doi:, http://www.sbs.utexas.edu/genetics/Literature/articles/NatureCellDivisionMilestones/ToCycleOrNot-Cancer.pdf
- ↑ a b Shiloh, Yosef (2003), "ATM and related protein kinases: safeguarding genome integrity", Nature Reviews Cancer 3 (3): 155–168, doi:, http://web.mit.edu/beh.109/www/Module3/handouts/Handout%20Lect%203%20shilo%20atm%20review%202003.pdf
- ↑ a b Goodarzi, A.A.; Block, W.D.; Lees-miller, S.P. (2003), Cell Cycle Res 5: 393–411, http://www.google.co.uk/webhp?hl=en
- ↑ Venkitaraman, A.R. (2001), "Functions of BRCA1 and BRCA2 in the biological response to DNA damage", Journal of Cell Science 114 (20): 3591–3598, http://jcs.biologists.org/cgi/content/full/114/20/3591
- ↑ Burke D.J., Stukenberg P.T. (2008). «Linking Kinetochore-Microtubule Binding to the Spindle Checkpoint.» Developmental Cell. Vol. 14. n.º 4. 474-479. [3]
Categoría: Ciclo celular

Wikimedia foundation. 2010.