- Virus de la inmunodeficiencia humana
-
 Virus de la inmunodeficiencia humana (VIH)
Virus de la inmunodeficiencia humana (VIH)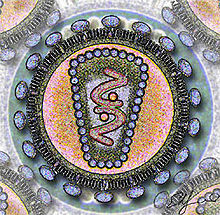
Corte del virus de la inmunodeficiencia humanaClasificación de los virus Grupo: VI (Virus ARN monocatenario retrotranscrito) Familia: Retroviridae Género: Lentivirus Especies - VIH Tipo 1
- VIH Tipo 2
El virus de la inmunodeficiencia humana (VIH) es un lentivirus (de la familia Retroviridae). Fue descubierto y considerado como el agente de la naciente epidemia de sida por el equipo de Luc Montagnier en Francia en 1983. El virión es esférico, dotado de una envoltura y con una cápside proteica. Su genoma es una cadena de ARN monocatenario que debe copiarse provisionalmente al ADN para poder multiplicarse e integrarse en el genoma de la célula que infecta. Los antígenos proteicos de la envoltura exterior se acoplan de forma específica con proteínas de la membrana de las células infectables, especialmente de los linfocitos T CD4.
El proceso de conversión de ARN en ADN es una característica principal de los retrovirus y se lleva a cabo mediante acciones enzimáticas de transcriptasa inversa. Con la demostración de la existencia de la transcriptasa inversa, se inició en la década de 1970 la búsqueda de los retrovirus humanos, que permitió el aislamiento en 1980 del virus de la leucemia de células T del adulto, HTLV-I (R. Gallo y cols.)
El VIH tiene un diámetro de aproximadamente 100 nanómetros. Su parte exterior es la "cubierta", una membrana que originalmente pertenecía a la célula de donde el virus emergió. En la cubierta se encuentra una proteína del virus, la gp41, o "glicoproteína transmembrana". Conectada a la gp41 está la gp120, la cual puede unirse al receptor CD4 localizado en la superficie de los linfocitos T para penetrar en ellos. El núcleo tiene la "cápside", compuesta por la proteína p24. En su interior está el ARN, la forma de información genética del VIH.
En diciembre de 2006, de acuerdo con la Organización Mundial de la Salud, había 39,5 millones de personas con VIH en el mundo, de las cuales 24,7 millones vivían en África Subsahariana.
Clasificación
El virus de inmunodeficiencia humana forma parte del género Lentivirus.[1] Estos constituyen un grupo dentro de la familia Retroviridae.[2] Los virus de este grupo poseen propiedades morfológicas y biológicas comunes. Varias especies son atacadas por los lentivirus, cuya característica principal consiste en un período de incubación prolongado que desemboca en enfermedad después de varios años.[3]
Desde su ingreso a la célula hospedadora, la cadena simple de ácido ribonucleico (ARN) viral comienza su transformación en una doble cadena de ácido desoxirribonucleico (ADN) por acción de la enzima transcriptasa inversa que forma parte del virus. La integrasa y otros cofactores actúan para que el ADN del virus se fusione con el ADN de la célula hospedadora[4] a través de la transcripción en el genoma de la célula que aloja al virus. De esta manera, la célula queda infectada por el virus. Después de este proceso, los lentivirus reaccionan de dos maneras posibles: puede ocurrir que el virus entre en latencia mientras la célula infectada continúa en funciones, o bien, que el virus comience a replicarse activamente y libere viriones capaces de infectar otras células.
Existen dos tipos del VIH, llamados VIH-1 y VIH-2. El primero de ellos corresponde al virus descubierto originalmente, que recibió los nombres de LAV y HTLV-III por parte de los dos equipos que estaban investigando el agente etiológico del sida durante la primera mitad de la década de 1980. El VIH-1 es más virulento e infeccioso que el VIH-2[5] y es el causante de la mayoría de infecciones por VIH en el mundo. El VIH-2 es menos contagioso y por ello se encuentra confinado casi exclusivamente a los países de África occidental.[6]
Estructura y genoma del VIH
Estructura
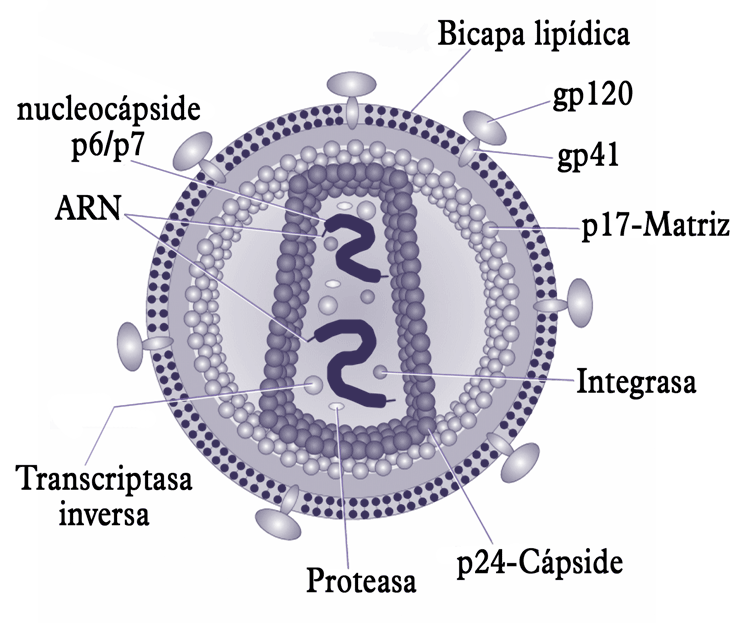
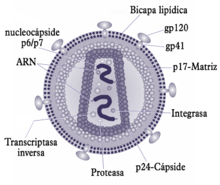 Estructura del VIH.
Estructura del VIH.
El VIH comparte con los retrovirus las características esenciales de esa familia. El virión contiene información genética bajo la forma de ácido ribonucléico (ARN), protegido por una envoltura de membrana. Los retrovirus insertan su información genética en las células hospedadora por acción de la transcriptasa inversa.[7]
Un virión del VIH tiene una forma aproximadamente esférica con un diámetro de 80-100 nm. Está constituido por tres capas. La exterior es una bicapa lipídica. Posee 72 prolongaciones formadas por las glicoproteínas gp120 y gp41 que actúan en el momento de la unión del virus a la célula hospedadora. La capa intermedia está constituida por la nucleocápside icosaédrica. la capa interior tiene forma de un cono truncado. Está constituida por el ARN viral y la nucleoproteína. La cadena genética del VIH está constituida por un ARN de cadena simple compuesto por dos filamentos idénticos. El ARN contiene varios genes, cada uno de los cuales codifica las diversas proteínas que el VIH necesita para reproducirse.[8]
Genoma y composición
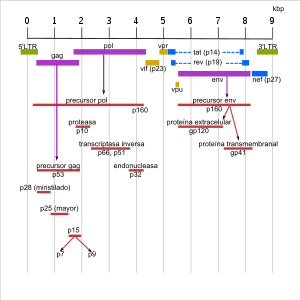 Genoma del VIH-1.
Genoma del VIH-1.Los genomas del VIH-1 y VIH-2 son muy similares. Ambos están compuestos por los tres genes básicos de la familia de los retrovirus. Se trata de los genes gag, pol y env. Cada uno de estos genes codifica proteínas que ayudan a la reproducción del virus. El genoma del VIH posee otros seis genes adicionales: tat, rev, vpu (vpx en el caso del VIH-2), vif y nef.[9]
Genes estructurales
Las proteínas estructurales son codificadas por los genes gag, pol y env, y su secuencia cubre la mayor parte del genoma viral, quedando sólo una parte menor para el resto de los genes.
El gen gag es traducido a una proteína precursora, la p55, que luego se asocia, durante la gemación por la que se liberan nuevas partículas víricas desde de la célula infectada, a dos copias del ARN viral, para el que presenta una región afín, y a otras proteínas virales y celulares. Una proteasa, producto del gen pol corta durante la maduración del virión la p55 en cuatro proteínas que se incorporan a sus lugares respectivos:
- La proteína p24 forma la cápside.
- La proteína p17 constituye la matriz, situada bajo la envoltura, a la que estabiliza. Una parte de las proteínas se unen al complejo molecular que acompaña al ADN viral al interior del núcleo. En la superficie de la proteína existe una región cariofílica (literalmente afin al núcleo) que es reconocida por la maquinaria molecular de importación nuclear. Éste es el mecanismo que permite al VIH infectar células diferenciadas, no destinadas a dividirse, algo que no ocurre en ningún otro retrovirus.
- Las proteínas p6 y p7 (ó p9) forman la nucleocápside. La región de la p55 correspondiente al polipéptido p6 es responsable de la incorporación de la proteína accesoria Vpr (producto de la traducción del gen vpr) al virión en formación y de la interacción con la membrana de la célula que hace posible la gemación. La p7 (p9) es responsable del reconocimiento y la incorporación del ARN al virión y además interviene en la transcripción inversa facilitándola.
Dentro de la cápside, además de las dos copias idénticas del ARN viral hay ejemplares de tres enzimas necesarias para la multiplicación del virus: una transcriptasa inversa, una integrasa y una proteasa. Estas enzimas, así como una ARNasa se producen a partir de la proteína Pol, después del corte de una proteína precursora mixta derivada de la cotraducción, una de cada 20 veces, de los genes gag y pol. La propia proteasa vírica rompe la proteína anterior, con una eficiencia limitada, para obtener las proteínas Gag (p55) y Pol. Luego la proteína precursora Pol es cortada a su vez para formar las cuatro proteínas funcionales citadas:
- La proteasa (p10). Se trata de una aspartil-proteasa cuya forma funcional es un dímero del que se conoce la estructura tridimensional. Actúa cortando las piezas de las proteínas Gag, Pol y de la Gag-Pol. Una parte de los fármacos empleados contra el VIH son inhibidores de su función.
- La transcriptasa inversa (p50) cuya función es la síntesis del ADN de doble cadena del provirus usando como patrón la cadena singular del ARN viral. Es una ADN-polimerasa que puede actuar como dependiente del ADN tanto como del ARN. Una vez formada la primera cadena de ADN, complementaria del ARN viral, la ARNasa lo separa de él, lo que permite a la transcriptasa inversa ejecutar la síntesis de la segunda cadena de ADN tomando como molde la primera que se formó. Así pues, para la síntesis de la primera cadena la actividad de la transcriptasa inversa es ARN-dependiente, pero para la de la segunda es ADN-dependiente. También existen múltiples fármacos contra la actividad de la transcriptasa inversa.
- La ARNasa (p15), que como se ha dicho separa las cadenas de ARN de las de la ADN durante la transcripción inversa.
- La integrasa (p31) realiza la inserción del ADN proviral en el genoma de la célula huésped. No se requiere ATP para su actividad y debe cumplir sucesivamente tres funciones:
- Con una actividad exonucleasa corta dos núcleótidos del extremo 3' de cada una de las dos cadenas del ADN proviral.
- Con una actividad endonucleasa (de doble cadena) corta el ADN del huésped en el punto de integración. No hay un lugar fijo en el genoma para que esto se realice, sino que ocurre en cualquier región muy accesible de la cromatina, lo que se supone que favorece la expresión del provirus, al coincidir esas regiones del genoma con las más transcritas.
- Por último, con una actividad ligasa el ADN proviral es soldado, mediante sólo un enlace covalente en cada extremo, en el ADN celular.
- En la actualidad existe un fármaco comercializado contra la actividad de la integrasa, el raltegravir.
La envoltura se basa en una bicapa lipídica, lo mismo que cualquier membrana biológica, y sus componentes estructurales básicos proceden de la membrana plasmática de la célula parasitada. Pero la envoltura porta además regularmente espaciadas 72 espículas, que son complejos proteicos integrados en la membrana formados por proteínas virales codificadas por el gen env. Cada espícula está formada por una pieza de la proteína gp41, integral en la membrana, y una cabeza externa formada por la proteína gp120, esencial para el acoplamiento con el exterior de ciertas células previo a su invasión. Entre los dos componentes de las espículas existe una unión no covalente. Las proteínas gp41 y gp120 se sintetizan como una sola poliproteína, gp160, con la información del gen env antes de que sea cortada por una proteasa de la célula. La proteína Env existe como trímero en la superficie de los viriones y las células infectadas.
Los fármacos inhibidores de la fusión funcionan contra la proteína gp41, para evitar su unión a los linfocitos.
Proteínas reguladoras
Tat
La proteína Tat existe en dos formas, una larga, de 101 restos aminoácidos de longitud, y otra más corta, de sólo 72. La segunda se produce cuando en fase temprana se produce una edición completa del ARNm viral, la primera cuando en una fase más tardía sólo se realiza una edición parcial. La proteína Tat (por transactivator) es imprescindible para la producción de nuevos viriones, que promueve activamente. La proteína se une a una región de 59 nucleótidos situada en el extremo 5' del ARN viral llamada TAR (Transactivator Active Region) y actúa como un transactivador, algo excepcional, puesto que éstos suelen unirse al ADN, no al ARN. En cuanto este extremo inicial del genoma viral ha sido transcrito desde el ADN proviral, la proteína Tat se une a él y promueve su elongación favoreciendo la transcripción del resto de la cadena.
Rev
La proteína rev regula la expresión del ARN viral controlando el ritmo de exportación del ARNm.[10]
Tat y Rev: acción conjunta
La acción sinergística de Tat y Rev fuertemente incrementa la expresión de proteínas virales. Los papeles que Tat y Rev desempeñan en la regulación transcripcional del VIH-1 y en la expresión de proteínas estructurales, respectivamente, hacen Tat y Rev esenciales para el ciclo de vida del VIH. Sus funciones facilitan la expresión de proteínas virales en dos etapas. Después de la integración del ADN proviral y de su transcripción en un nivel basal, solamente los RNAms de 2 KB se transportan al citoplasma. Esto permite la síntesis de Tat, Rev y de Nef. Tat y Rev entonces son transportadas al núcleo, donde actúan para aumentar la transcripción del ADN del provirus (Tat) y del transporte de todos los RNAms virales al citoplasma (Rev). La expresión de proteínas codificada por las clases de RNAm de 9 KB y 4 KB (Gag, Gag-Pol, Env, Vpr, Vif, y de Vpu) puede entonces ocurrir. Estudios donde se han mutado genes virales han determinado que Vif, Vpr, Vpu y Nef no son esenciales para la producción de partículas infecciosas en cultivos celulares in-vitro. Sin embargo, la conservación de dichas proteínas accesorias en el genoma del VIH sugiere que todas desempeñan papeles importantes durante el ciclo infeccioso en el huésped. Los roles de estas proteínas serán descritos a continuación.
Proteínas accesorias
Vif: incremento en infectividad y protección del genoma viral
Vif es una proteína de 193 aminoácidos que está presente en bajos niveles adentro de los viriones, e interactúa con en RNA genómico viral. La división de esta proteína reduce la infectividad del VIH-1 en cultivos celulares y en modelos animales de patogénesis. No obstante, el mecanismo de acción de Vif se ha empezado a entender recientemente. La ausencia de Vif en partículas infecciosas no puede ser compensada con la expresión de Vif en las células infectadas. Estudios recientes han demostrado que Vif es requerida para eliminar la acción del factor ApoBEC3G, la cual es una deaminasa de citidinas, que convierte la citosina en uracilo, y emplea como sustrato el ADN de cadena sencilla. Además, esta enzima posiblemente actúa durante el ciclo de la transcriptasa inversa, modificando así la cadena negativa del DNA, porque esta es la fase en la cual el ADN de cadena sencilla está disponible. ApoBEC3G es selectivamente incorporada dentro de las partículas de VIH, resultando en un alto nivel de mutaciones en el genoma viral. Dado que estos altos niveles de mutación son perjudiciales para la viabilidad del virus, VIH ha evolucionado una estrategia para abolir esta poderosa barrera. Sin embargo, estudios recientes sugieren que ApoBEC3G no requiere su acción enzimática para tener efecto. Estudios más recientes han implicado que ApoBEC3G tiene un rol en la inhibición de ciertas fases en el ciclo de la transcriptasa inversa.
Vpu: facilita el desprendimiento de viriones en células infectadas
Vpu es una proteína de 81 aminoácidos que es insertada en membranas vía su terminal nitrogenado. Vpu se acumula en el aparato de Golgi y en endosomas celulares. Vpu es única en HIV-1 y no hay homólogos en lentiviruses relacionados como el VIH-2 y el VIS. A Vpu se le han atribuido dos actividades.
Degradación de la proteína CD4
En la ausencia de Vpu, la proteína CD4 interactúa con la proteína viral gp160 recién sintetizada para formar un complejo insoluble, el cual retiene gp120 dentro de la célula. La región citoplásmica de Vpu se puede unir con CD4 y con la proteína β-TrCP. Esto induce la ubiquitinizacion de CD4 y su subsiguiente degradación por el proteasoma, incrementando así la expresión de gp120 en la superficie celular.
Realza en el desprendimiento de viriones de la membrana celular
Esta actividad depende de la región transmembranal de Vpu. En la ausencia de Vpu, los viriones se acumulan en la superficie celular en un estado parcialmente desprendido. Expresión de Vpu resulta en la liberación facilitada de viriones de la membrana celular. Remarcablemente, este efecto no está restringido solamente al VIH-1; Vpu también facilita el desprendimiento de otros virus no relacionados. El mecanismo por la cual esto ocurre es desconocido. Se ha sugerido que Vpu facilita la fluidez de la membrana celular por medio de un canal de cationes. También se ha sugerido que Vpu causa disrupción de interacciones entre proteínas del VIH y de la superficie celular; esto previene la endocitosis de viriones recientemente desprendidos de la célula...
Ciclo de replicación
Las células que el VIH invade son esencialmente los linfocitos T CD4+, pero también en menor medida los monocitos/macrófagos, las células dendríticas, las células de Langerhans y las células de microglía del cerebro. La replicación viral tiene pues lugar en tejidos diversos (de ganglios linfáticos, intestino, cerebro, timo,…). Los órganos linfoides, sobre todo los ganglios linfáticos, constituyen la principal sede de su replicación. El virus está presente en numerosos líquidos del organismo, en particular la sangre y las secreciones genitales.
La replicación del virus se desarrolla en las siguientes etapas:
La fijación representa la primera etapa en la invasión de una célula. Se basa en el reconocimiento mutuo y acoplamiento de proteínas de la envoltura del virión, las gp120 y gp41, y los receptores de la célula blanca, los CD4. Este reconocimiento no es posible sin ayuda de correceptores propios de las células susceptibles de ser invadidas; en el caso de los macrófagos son los CCR5 y en el caso de los LT4, los CXCR4, que interactúan con la proteína superficial. Macrófagos y LT4 tienen en común su principal receptor: el receptor CD4. Este reconocimiento es condición obligada para que el virus llegue a penetrar en la célula y continuar con el proceso de infección. La penetración es el segundo paso: una vez reconocido el virión por los receptores de superficie, se vacía dentro de la célula fusionándose la envoltura lipídica del virión con la membrana plasmática de la célula. Protegidos por la cápside y las nucleocápsides, los dos ARN mensajeros que forman el genoma viral y sus proteínas asociadas se encuentran ahora en el citoplasma. Eliminación de las cubiertas proteicas, cápside y nucleocápsides, quedando el ARN vírico libre en el citoplasma y listo para ser procesado. La transcripción inversa del ARN vírico para formar ADNc (ADN complementario, monocatenario) con la misma información. Cada una de las dos moléculas de ARN llega desde el virión asociada a una molécula de transcriptasa inversa que se ocupa del proceso. Las dos moléculas de ADNc se asocian para formar una molécula de ADN, que es la forma química de guardar la información que una célula eucariota es capaz de procesar. El paso siguiente es la integración del genoma vírico en el genoma de la célula huésped. Para ello penetra en el núcleo y se inserta en el ADN celular con ayuda de una integrasa, que procede del virión infectante. La transcripción del ADN vírico por los mecanismos normales de la célula. El resultado de la transcripción es un ARNm (ARN mensajero). El ARNm obtenido es complejo, constituido por una sucesión de intrones (partes no informativas) y exones (partes informativas). Debe ser procesado por cortes y reempalmes antes de que la información que contiene pueda servir para fabricar las proteínas correspondientes. Una vez procesado, el ARNm puede salir del núcleo a través de los poros nucleares. Una vez en el citoplasma el ARNm proporciona la información para la traducción, es decir, la síntesis de proteínas, que es realizada a través del aparato molecular correspondiente, del que forman la parte fundamental los ribosomas. El resultado de la traducción no consiste inmediatamente en proteínas funcionales, sino en poliproteínas que aún deben ser cortadas en fragmentos. Por acción de proteasas específicas del VIH, las poliproteínas producto de la traducción son procesadas, cortándolas, para formar las proteínas constitutivas del virus. Las proteínas víricas fabricadas se ensamblan, junto con ARN provirales, para formar los componentes internos de la estructura del virión, los que constituyen la cápside y su contenido. El último paso es la gemación, cuando los nucleoides víricos se aproximan a la membrana plasmática y se hacen envolver en una verruga que termina por desprenderse, formando un nuevo virión o partícula infectante. En cada célula infectada se ensamblan varios miles de nuevos viriones, aunque muchos son incompletos y no pueden infectar.
Mecanismos de transmisión del virus
Riesgo estimado de adquisición del VIH
según el tipo de exposición[11]Tipo de exposición Número estimado de infecciones por cada 10.000 exposiciones a una fuente infectada Transfusión de sangre 9,000[12] Parto 2,500[13] Inyección de droga- 67[14] Coito anal receptivo* 50[15] [16] Aguja de laboratorio percutánea 30[17] Coito vaginal receptivo* 10[15] [16] [18] Coito anal insertivo* 6.5[15] [16] Coito vaginal insertivo* 5[15] [16] Felación receptiva* 1[16] Felación insertiva* 0.5[16] * sin uso de preservativo El VIH sólo se puede transmitir a través del contacto entre fluidos corporales que poseen una alta concentración viral. El virus no se transmite de manera casual. De acuerdo con los CDC de Estados Unidos, no se han encontrado casos en que abrazos, besos secos o saludos con las manos hayan sido causantes de infección.[19] El virus ha sido aislado en la saliva, las lágrimas y la orina, el semen, el líquido preseminal, los fluidos vaginales, el líquido amniótico, la leche materna, el líquido cefalorraquídeo y la sangre, entre otros fluidos corporales humanos.
Las tres principales formas de transmisión son:
- Sexual (acto sexual sin protección). (infección de transmisión sexual). La transmisión se produce por el contacto de secreciones infectadas con la mucosa genital, rectal u oral de la otra persona.
- Parenteral (por sangre). Es una forma de transmisión a través de jeringuillas contaminadas que se da por la utilización de drogas intravenosas o a través de los servicios sanitarios, como ha ocurrido a veces en países pobres, no usan las mejores medidas de higiene; también en personas, como hemofílicos, que han recibido una transfusión de sangre contaminada o productos contaminados derivados de la sangre; y en menor grado trabajadores de salud que estén expuestos a la infección en un accidente de trabajo como puede ocurrir si una herida entra en contacto con sangre contaminada; también durante la realización de piercings, tatuajes y escarificaciones.
- Vertical (de madre a hijo). La transmisión puede ocurrir durante las últimas semanas del embarazo, durante el parto, o al amamantar al bebé. De estas situaciones, el parto es la más problemática. Actualmente en países desarrollados la transmisión vertical del VIH está totalmente controlada (siempre que la madre sepa que es portadora del virus) ya que desde el inicio del embarazo (y en ciertos casos con anterioridad incluso) se le da a la embarazada un Tratamiento Anti-Retroviral de Gran Actividad (TARGA) especialmente indicado para estas situaciones, el parto se realiza por cesárea generalmente, se suprime la producción de leche, y con ello la lactancia, e incluso se da tratamiento antiviral al recién nacido.
Profilaxis de emergencia
Si una persona contrae el VIH debido a alguna circunstancia imprevista (la penetración de una aguja en un laboratorio, una violación o un condón que se rompe durante el coito), puede aplicarse entonces lo que se conoce como tratamiento profilaxis post-exposición para el VIH. Este es un régimen de medicamentos muy potentes contra el VIH que pueden aplicarse en la hora siguiente al incidente y que siguen ejerciendo su efecto durante las primeras 72 horas (su eficacia va disminuyendo con cada hora transcurrida desde el evento). Este tratamiento puede evitar que la persona se vuelva seropositiva al VIH.
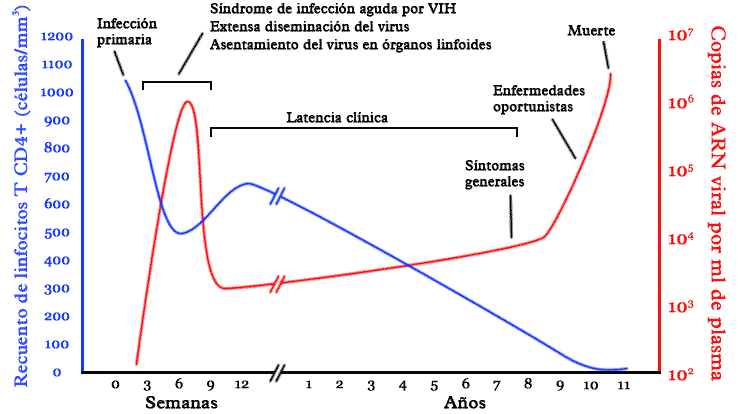
Historia natural de la infección por VIH
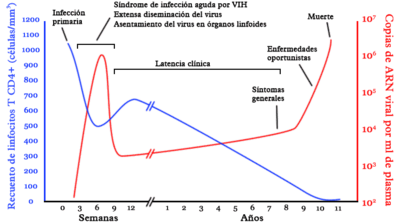 Diagrama sobre la historia natural de la infección por VIH.
Diagrama sobre la historia natural de la infección por VIH.La infección por VIH se presenta en diversas etapas, identificadas por un conjunto de síntomas e indicadores clínicos. En ausencia de un tratamiento adecuado, el virus se replica constantemente e infecta los linfocitos T-CD4, que constituyen una parte esencial del sistema inmunológico en los seres humanos. Por su parte, el sistema inmunológico del portador del VIH reacciona ante la presencia del virus y genera una respuesta que puede mantener la infección bajo control al menos por un tiempo, mediante la reposición de células defensivas. Al término de un período que se puede prolongar por varios años, el VIH se vuelve resistente a las defensas naturales del cuerpo y destruye el sistema inmune del portador. De esta manera, la persona seropositivo queda expuesta a diversas enfermedades oportunistas y fallece.[20]
Fase aguda
La fase de la infección aguda por VIH inicia en el momento del contagio. El virus se propaga por el cuerpo de la persona contagiada a través de sus fluidos corporales. En un plazo de días, el VIH infecta no sólo las células expuestas inicialmente (por ejemplo, las células de la mucosa vaginal o rectal en el caso de una infección por vía sexual)[21] sino también los ganglios linfáticos. Durante ese tiempo, el VIH se multiplica dentro del organismo hasta alcanzar niveles propios de la infección crónica. El tejido linfoide asociado a los intestinos constituye uno de los principales espacios del cuerpo humano donde tiene lugar la reproducción inicial del VIH por su alto porcentaje de linfocitos T CD4.[22]
Un porcentaje importante de personas que contraen el virus no presenta síntomas de la infección en su fase aguda. Es decir, son pacientes asintomáticos. Sin embargo, se calcula que entre el 40/50%-90% o hasta el 80%[23] de los casos de contagio con VIH-1 presentan manifestaciones clínicas. El cuadro de la infección aguda es similar al de una mononucleosis infecciosa: fiebre, malestares musculares, inflamación de los ganglios, sudoración nocturna, diarrea, náuseas y vómito. La gran mayoría de los seropositivos no reciben diagnóstico del cuadro agudo de la infección por VIH, pues son síntomas compartidos por varias enfermedades. Por lo tanto, presentar un conjunto de síntomas como el descrito aquí no es indicador necesario de que una persona se haya infectado por VIH, aunque es recomendable que quien considere que ha estado expuesto al contagio y presente los síntomas, acuda a un especialista para recibir atención médica.[24] El cuadro de la infección aguda por VIH aparece entre dos y seis semanas después de la exposición al virus, y desaparece unos pocos días después.[25]
El VIH ataca principalmente los linfocitos T CD4+, que forman parte del sistema inmune de los seres humanos. Aunque estas células por sí mismas no tienen una función de ataque contra células extrañas al cuerpo, tienen un papel importante en la respuesta inmunológica adaptativa. En una persona con buena salud, el número de linfocitos T CD4+ oscila entre 1200 y 500/μl. Durante la fase asintomática de la infección, la proporción de linfocitos infectados 1/1000-1/100 000, que aumentará progresivamente hasta llegar a 1/100 en la infección crónica.[26] Durante la fase aguda de la infección, las pruebas tradicionales siempre darán negativo porque no detectan directamente el VIH, sino los anticuerpos producidos como respuesta por el sistema inmune, lo que ocurre alrededor de la 12a semana después de la exposición. En contraste, las pruebas de carga viral, que contabilizan el número de copias del ARN del virus en la sangre, arrojarán como resultado una elevada cantidad de copias del VIH durante la fase aguda de la infección.[27]
Fase crónica
La fase crónica de la infección por VIH se suele llamar también latencia clínica porque el portador es asintomático, es decir, no presenta síntomas que puedan asociarse con la infección.[28] Esto no quiere decir que el virus se encuentre inactivo. Por el contrario, durante la fase crónica el VIH se multiplica incesantemente. Se calcula que, en un sujeto infectado, diariamente se producen entre mil y diez mil millones de nuevas partículas virales y son destruidos alrededor de cien millones de linfocitos T CD4.[29] Los pacientes son asintomáticos gracias a que el sistema inmune tiene una gran capacidad para regenerar las células destruidas por el virus, pero pueden presentar adenopatías y la disminución del conteo de plaquetas en la sangre.
La reacción ante la presencia del virus termina por desgastar al sistema inmunológico. En ausencia de tratamiento, la mayoría de los portadores del virus desarrollan el síndrome de inmunodeficiencia adquirida (sida) en un plazo de 5 a 10 años. La causa de esto es que, mientras el virus sigue reproduciéndose de manera constante y aumenta la carga viral en su anfitrión, disminuye también la capacidad de recuperación del sistema inmune. Al término fase crónica, los pacientes desarrollan otras manifestaciones de la infección como dermatitis seborréica, úlceras bucales y foliculitis.[30]
Síndrome de inmunodeficiencia adquirida
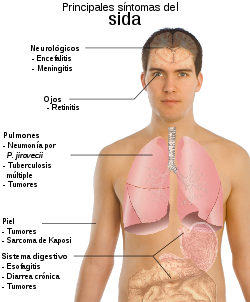 Principales síntomas del sida
Principales síntomas del sidaEl sida constituye la etapa crítica de la infección por VIH. En esta fase de la infección, el portador del VIH posee un sistema inmunológico que probablemente sea incapaz de reponer los linfocitos T CD4+ que pierde bajo el ataque del VIH y también ha visto reducida su capacidad citotóxica hacia el virus. Este fenómeno coincide con el aumento en las tasas de replicación del virus, que merma la capacidad de reacción del anfitrión ante otros agentes causantes de enfermedades. De esta manera, el portador del virus es presa potencial de numerosas infecciones oportunistas que le pueden conducir a la muerte. La neumonía por P. jiroveci, el sarcoma de Kaposi, la tuberculosis, la candidiasis y la infección por citomegalovirus son algunas de las infecciones más frecuentes que atacan a los seropositivos que han desarrollado sida.[31]
La mayoría de los pacientes que han desarrollado sida no sobreviven más de tres años sin recibir tratamiento antirretroviral. Sin embargo, incluso en esta fase crítica el sida y el VIH pueden ser controlados mediante la terapia antirretroviral de gran actividad. Los antirretrovirales pueden brindar una mejor calidad de vida a un portador del VIH y aumentan sus posibilidades de supervivencia. Dado que el VIH tiene una gran capacidad de mutación, con el tiempo los antirretrovirales pierden su efectividad porque el virus desarrolla resistencia a ellos. Una vez que esto ocurre, el paciente queda expuesto nuevamente a las infecciones oportunistas y, eventualmente, a la muerte, en tanto que no se dispone de un medicamento que cure la infección por VIH.
Historia
Origen y evolución
 El VIH-1 está relacionado con el SIVcpz que ataca a los chimpancés.
El VIH-1 está relacionado con el SIVcpz que ataca a los chimpancés.Como otros agentes causantes de enfermedades infecciosas emergentes, el VIH pasó a los seres humanos por zoonosis, es decir por contagio desde otras especies. La emergencia del sida y la identificación del VIH estimularon investigaciones que han permitido determinar que las variantes del VIH forman parte de un amplio grupo de lentivirus. El VIH es sumamente parecido a un virus que ataca a los primates. Se trata del virus de inmunodeficiencia de los simios (Simian immunodeficiency virus, SIV), del que se conocen diversas cepas se transmiten por vía sexual.[32] A diferencia del VIH, el virus de los primates no causa inmunodeficiencia en los organismos que lo hospedan, salvo en el caso del salto de una especie a otra.
El VIH-1, responsable de la actual pandemia, ha resultado estar estrechamente relacionado con el SIVcpz, que infecta a poblaciones de la subespecie centroafricana del chimpancé común (Pan troglodytes troglodytes). El SIVcpz, a su vez, parece derivar por recombinación (un fenómeno que se produce fácilmente cuando infectan al mismo individuo dos cepas víricas diferentes) del SIVrcm, propio del mangabeye de collar (Cercocebus torquatus), y del SIVgsn, propio del avoem (Cercopithecus nictitans)[33] Esta hipótesis es sostenida por el hecho de que tanto el VIH como las diversas cepas del SIV poseen el gen vpu, además de que se han reportado contagios por SIV entre humanos en África ecuatorial.[34] Las distribuciones actuales de las especies implicadas se solapan, y de los chimpancés se sabe que cazan monos pequeños para comerlos, lo que habría facilitado la coinfección por cepas diversas de SIV. La subespecie oriental del chimpancé, Pan troglodytes schweinfurthi, presenta también infección con una cepa propia del SIVcpz, pero genéticamente alejada del clado formado por el VIH-1 y las cepas de P.t.troglodytes. No se ha encontrado presencia del SIVcpz en la subespecie occidental, P. t. verus, aunque se observó el contagio en cautividad de un individuo de esta subespecie.
El salto de la barrera de especie desde P. t. troglodytes a Homo sapiens sapiens se ha producido al menos tres veces, con variantes del VIH-1 que demuestran parentesco con distintas cepas, geográficamente más o menos localizadas, del SIVcpz. Así pues, el VIH-1 es un virus polifilético. El grupo M del VIH-1, responsable de la pandemia actual, debió pasar a los seres humanos en la primera mitad del siglo XX. Los grupos O y N del VIH-1 están restringidos a África Occidental ecuatorial, con el grupo N presente sólo en Camerún. Con los datos actuales, parece claro que Pan troglodytes troglodytes es el reservorio desde el que se han producido repetidamente las infecciones humanas por los virus de cuya evolución procede el VIH-1[32]
A su vez el VIH-2, extendido en África Occidental, procede del SIVsm, propio del mangabeye fuliginoso (Cercocebus atys atys), que habita las selvas costeras desde Senegal hasta Costa de Marfil. El análisis filogenético muestra que el paso a los seres humanos ha ocurrido también varias veces.[35]
Los SIV identificados hasta ahora se encuentran, de forma específica, en unas 35 especies de primates africanos, aproximadamente la mitad de las 70 que existen al sur del Sahara, y es en África donde parece tener su origen evolutivo este grupo monofilético de virus, genéticamente bien delimitado del resto de los lentivirus. La prevalencia (frecuencia de la infección) es variable entre especies y poblaciones, aunque no superior al 30%, en las poblaciones afectadas de chimpancés, pero puede pasar del 50% en poblaciones de otros primates, como Cercocebus atys.
En todos los casos conocidos el virus parece encontrarse cerca del equilibrio con su huésped natural, como resultado probable de una más o menos larga coevolución, observándose generalmente sólo versiones muy atenuadas del síndrome de inmunodeficiencia, como una reducción limitada de linfocitos T CD4+, reducción que no compromete en general la vida del individuo, aunque en un ejemplar de Cercocebus atys se produjo un sida típico después de 18 años de incubación. Este dato hace pensar que, al menos en parte, es la baja longevidad, unida a una larga incubación, lo que hace que la inmunodeficiencia sobrevenida sea un resultado excepcional de la infección en monos[36]
Descubrimiento
 De izquierda a derecha en la foto, Montagnier, Barré-Sinoussi y zur Hausen tras recibir el Premio Nobel de Medicina en 2008. Los dos primeros fueron reconocidos por el descubrimiento del VIH.
De izquierda a derecha en la foto, Montagnier, Barré-Sinoussi y zur Hausen tras recibir el Premio Nobel de Medicina en 2008. Los dos primeros fueron reconocidos por el descubrimiento del VIH.Desde 1981 se detectaron casos sorprendentes de infección por Pneumocystis jiroveci (entonces designado Pneumocystis carinii), un hongo emparentado con las formas originales de los Ascomycetes, conocido por infectar a pacientes severamente inmunodeprimidos. Inicialmente se observó un grupo de casos semejantes en los que estaban implicados varones homosexuales y donde aparecían a la vez infección por citomegalovirus y candidiasis. Se pensó primero que la causa debía estar ligada a prácticas comunes entre la población homosexual masculina.
Pronto empezaron a aparecer casos que afectaban a varones o mujeres heterosexuales usuarios de drogas intravenosas, así como a sus hijos; también entre pacientes no homosexuales y con hábitos saludables que habían recibido transfusiones de sangre entera o de productos sanguíneos por su condición de hemofílicos. Pronto se pensó, por criterios básicamente epidemiológicos, que la causa debía ser un agente infeccioso que se transmitía de forma semejante a como lo hace el virus de la hepatitis B.
Distintos equipos empezaron a buscar un virus asociado a los casos conocidos de inmunodeficiencia adquirida, tal vez un retrovirus como el que se sabía producía la inmunodeficiencia del gato o como el HTLV, productor de un tipo de leucemia. En 1983, en el Instituto Pasteur de París, un equipo dedicado a la investigación de la relación entre retrovirus y cáncer dirigido por J.C. Chermann, F. Barré-Sinoussi y L. Montagnier, encontró un candidato al que denominó lymphadenopathy-associated virus (virus asociado a la linfoadenopatía, LAV).
En 1984 el equipo de R. Gallo, descubridor del HTLV, único retrovirus humano conocido entonces, confirmó el descubrimiento, pero llamando al virus human T lymphotropic virus type III (virus linfotrópico T humano tipo III, con las siglas HTLV-III). Se produjo una subsecuente disputa sobre la prioridad en la que quedó claro que Gallo había descrito el virus sólo después de haber recibido muestras de los franceses. Como parte de la resolución del conflicto, el virus adquirió su denominación definitiva, human immunodeficiency virus (HIV) que en castellano se expresa como virus de la inmunodeficiencia humana (VIH).
En el mismo año, 1983, en que se identificó el virus, diversos equipos empezaron a trabajar en la secuencia de su genoma, publicada a principios de 1985, y comenzó también la caracterización de sus proteínas.
Epidemiología
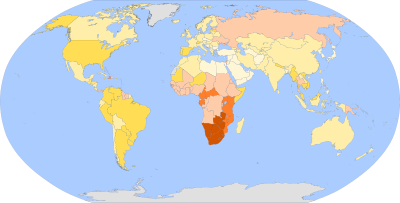 Prevalencia del VIH en el mundo (1982-1996). Clave: Sin datos Menos de 0.1% 0.1-0.5 % 0.5-1 % 1-5 % 5-15 % 15-50 %
Prevalencia del VIH en el mundo (1982-1996). Clave: Sin datos Menos de 0.1% 0.1-0.5 % 0.5-1 % 1-5 % 5-15 % 15-50 %El VIH se ha convertido en una epidemia de dimensiones mundiales. El Programa Conjunto de Naciones Unidas sobre el VIH/sida (Onusida) coordina esfuerzos internacionales de científicos, gobiernos, iniciativa privada y organizaciones civiles dirigidos a actuar sobre la epidemia del VIH y sus efectos.[37] Onusida observa el desarrollo epidemiológico de la infección por VIH en todo el mundo y emite un reporte sobre la situación de la epidemia cada dos años. Los informes de Onusida recopilan los datos provenientes de todos los países y dan una visión general de la evolución de la pandemia, sus efectos sociales, las estrategias adoptadas para controlarla.
Entre 1981 y 2007, el sida había causado la muerte de aproximadamente 25 millones de personas alrededor de todo el mundo.[38] En ese mismo año, 33 millones [30-36 millones] de personas estaban contagiadas con VIH. La epidemia se ha estabilizado en cuanto que no ha aumentado la proporción de personas infectadas respecto a la población total. Además se ha observado una reducción del total mundial de nuevos casos de infección por VIH, de 3 millones [2,6-3,5 millones] en 2002 a 2,7 millones [2,2-3,2 millones] en 2007.[39]
La región más afectada por la pandemia es África subsahariana, donde radican 21,5 millones [20,5-23,6 millones] de seropositivos. Esta cifra representa casi tres cuartos del total de casos calculados para todo el mundo. Esta región del mundo también presenta los índices más altos de mortalidad por sida y concentra el mayor número de nuevos contagios.[40]
Fármacos contra el VIH
Existen numerosos fármacos dirigidos a evitar tanto la infección, como la progresión del ciclo vital del virus. Dichos fármacos se clasifican según la proteína a la que van dirigidos (esto es, el paso replicativo que inhiben en su uso). En general, y dada la alta tasa de resistencias, está indicado el uso combinado de fármacos de diferentes grupos (politerapia), en lo que se viene llamando TARGA: Terapia AntirRetroviral de Gran Actividad.
Ninguno de estos fármacos ha mostrado ser efectivo por separado y, de hecho, uno de los más comunes, el llamado AZT, es altamente tóxico. El AZT por sí solo no puede destruir directamente el virus; lo que hace este fármaco es inhibir la enzima transcriptasa inversa, con lo que impide que el RNA del Virus se copie hacia cDNA bicatenario y, por consiguiente, evitar que se genere un provirus (el provirus es el cDNA que se integra al genoma de la célula huésped, en este caso es el linfocito T CD4+). Administrado de forma aislada, es decir, sin ser combinado con los otros medicamentos que componen el TARGA, puede incrementar las mutaciones en el virus que lo hagan más resistente y agresivo, anulando su eficacia terapéutica y acelerando el progreso de la enfermedad. Este riesgo disminuye notablemente cuando se combina con los otros medicamentos de la politerapia. También disminuye sensiblemente su toxicidad al reducirse y ajustarse con mejor precisión sus mínimas dosis efectivas en combinación con los otros componentes del TARGA.
Detección del VIH
Debido a que no existe ninguna manifestación clínica característica de la infección de VIH, la prueba para detectar esta enfermedad ha de llevarse a cabo mediante pruebas de diagnóstico molecular en un laboratorio. La prueba más habitual para detectar la presencia de VIH es la prueba de inmunodetección denominada ELISA. Con esta técnica se pretende detectar los anticuerpos específicos que el organismo produce como respuesta a la presencia del virus. Cabe destacar que, ante un resultado positivo mediante un ELISA, no se debe informar al paciente de la presencia de VIH sin haber confirmado antes la prueba mediante un western blot. Por lo tanto en la mayoría de los casos la seropositividad frente al VIH se detecta a partir de una extracción sanguínea del sujeto con la que se realizará la determinación de anticuerpos anti-VIH por alguna técnica de cribado como la ya nombrada ELISA u otras parecidas. La prueba diagnóstica dirigida al VIH tiene una especificidad del 99% y una sensibilidad del 99%.
Puntos de vista alternativos
Respecto a la existencia del VIH y respecto a su origen existen puntos de vista alternativos al consenso científico. Incluyen la teoría de E. Hooper de que la barrera de especie fue atravesada como consecuencia de la utilización de riñones infectados de chimpancé para producir vacunas contra la polio, o la opinión muy extendida entre jóvenes africanos de que el VIH es un arma biológica desarrollada por Estados Unidos contra los africanos, teoría que defienden debido a la extraña facilidad del virus en mutar de diferente forma en varios cuerpos infectados o en su misterioso origen aún no descubierto más de dos décadas después del primer caso de sida.
Existen numerosos activistas y algunos científicos que niegan que el sida sea causado por el VIH, algunos de los cuales incluso dudan de la misma existencia del virus aquí descrito. Muchos niegan también que exista el sida como entidad nosológica (una enfermedad singular bien definida) interpretando que la diversidad de formas epidemiológicas y clínicas es propia de una pluralidad de enfermedades que errónea o interesadamente son interpretadas como una sola. A este respecto véase Disidencia del VIH.
MVA-B: Perspectivas actuales de investigación
En la actualidad, un grupo de investigación español del CSIC, ha presentado una posible vacuna contra el VIH, la vacuna MVA-B.
La vacuna MVA-B se denomina así por su composición a partir del virus Vaccinia Modificado de Ankara (MVA), y la letra B procede del subtipo de VIH contra el que lucha, el más prevalente en Europa.
Esta vacuna se encuentra en la fase I de desarrollo y ha presentado una alta seguridad y eficacia. Un 90% de los voluntarios vacunados con MVA-B han generado una respuesta inmunitaria defensiva contra el VIH, y el 85% de ellos la ha mantenido, al menos durante un año.
Para el desarrollo de la vacuna MVA-B, se han introducido cuatro genes del VIH (Gag, Pol, Nef y Env) en la secuencia genética de vaccinia. Si el sistema inmune está sano reacciona frente al MVA, y los genes de VIH insertados en su secuencia no son capaces de infectar a los humanos, lo que garantiza la seguridad del ensayo clínico.[41] [42]
Pese a estos resultados esperanzadores, la vacuna todavía no puede ser comercializada, ya que ha de concluir con éxito todas las fases de desarrollo del ensayo clínico para poder salir al mercado.
Véase también
- CCR5
- complejo de demencia asociado al VIH
- infecciones de transmisión sexual
- negacionismo del VIH/sida
- pandemia de VIH/sida
- prueba de VIH
- retrovirus
- sida
- VIH en el niño
- VIH en la mujer
Referencias
Notas
- ↑ International Committee on Taxonomy of Viruses. «61.0.6. Lentivirus». National Institutes of Health. Consultado el 28-02-2006.
- ↑ International Committee on Taxonomy of Viruses. «61. Retroviridae». National Institutes of Health. Consultado el 28-02-2006.
- ↑ Lévy, J. A. (1993). «HIV pathogenesis and long-term survival». AIDS 7 (11asdd): pp. 1401–10. doi:. PMID 8280406.
- ↑ Smith, Johanna A.; Daniel, René (Division of Infectious Diseases, Center for Human Virology, Thomas Jefferson University, Philadelphia) (2006). «Following the path of the virus: the exploitation of host DNA repair mechanisms by retroviruses». ACS Chem Biol 1 (4): pp. 217–26. doi:. PMID 17163676.
- ↑ Gilbert, PB et al (28 de febrero de 2003). «Comparison of HIV-1 and HIV-2 infectivity from a prospective cohort study in Senegal». Statistics in Medicine 22 (4): pp. 573–593. doi:. PMID 12590415.
- ↑ Reeves, J. D. and Doms, R. W (2002). «Human Immunodeficiency Virus Type 2». J. Gen. Virol. 83 (Pt 6): pp. 1253–65. PMID 12029140.
- ↑ Garrido, Olmo y Castel, 2001: 321-322.
- ↑ Bravo, 2005: 685; Pascual y Corral, 2003: 29.
- ↑ Garrido, Olmo y Castel, 2001: 321.
- ↑ Importancia de la proteína Rev
- ↑ Smith, D. K., Grohskopf, L. A., Black, R. J., Auerbach, J. D., Veronese, F., Struble, K. A., Cheever, L., Johnson, M., Paxton, L. A., Onorato, I. A. and Greenberg, A. E. (2005). «Antiretroviral Postexposure Prophylaxis After Sexual, Injection-Drug Use, or Other Nonoccupational Exposure to HIV in the United States». MMWR 54 (RR02): pp. 1-20. http://www.cdc.gov/mmwr/preview/mmwrhtml/rr5402a1.htm.
- ↑ Donegan, E., Stuart, M., Niland, J. C., Sacks, H. S., Azen, S. P., Dietrich, S. L., Faucett, C., Fletcher, M. A., Kleinman, S. H., Operskalski, E. A., et al. (1990). «Infection with human immunodeficiency virus type 1 (HIV-1) among recipients of antibody-positive blood donations». Ann. Intern. Med. 113 (10): pp. 733-739. PMID 2240875.
- ↑ Coovadia, H. (2004). «Antiretroviral agents—how best to protect infants from HIV and save their mothers from AIDS». N. Engl. J. Med. 351 (3): pp. 289-292. PMID 15247337.
- ↑ Kaplan, E. H. and Heimer, R. (1995). «HIV incidence among New Haven needle exchange participants: updated estimates from syringe tracking and testing data». J. Acquir. Immune Defic. Syndr. Hum. Retrovirol. 10 (2): pp. 175-176. PMID 7552482.
- ↑ a b c d European Study Group on Heterosexual Transmission of HIV (1992). «Comparison of female to male and male to female transmission of HIV in 563 stable couples». BMJ. 304 (6830): pp. 809-813. PMID 1392708.
- ↑ a b c d e f Varghese, B., Maher, J. E., Peterman, T. A., Branson, B. M. and Steketee, R. W. (2002). «Reducing the risk of sexual HIV transmission: quantifying the per-act risk for HIV on the basis of choice of partner, sex act, and condom use». Sex. Transm. Dis. 29 (1): pp. 38-43. PMID 11773877.
- ↑ Bell, D. M. (1997). «Occupational risk of human immunodeficiency virus infection in healthcare workers: an overview.». Am. J. Med. 102 (5B): pp. 9-15. PMID 9845490.
- ↑ Leynaert, B., Downs, A. M. and de Vincenzi, I. (1998). «Heterosexual transmission of human immunodeficiency virus: variability of infectivity throughout the course of infection. European Study Group on Heterosexual Transmission of HIV». Am. J. Epidemiol. 148 (1): pp. 88-96. PMID 9663408.
- ↑ CDC (2006): "Preguntas y Respuestas: Transmisión", en The Body, 18 de octubre de 2006. Consultado el 10 de noviembre de 2010.
- ↑ Pérez, Bayón y Esquitino, 2003: 85-91.
- ↑ No está claro cuál es el tipo de células que son afectadas inicialmente por el VIH, ni se conocen con precisión los mecanismos a través de los cuales son infectadas las células de los tejidos linfoides después de la primoinfección (Dybull, Connors y Fauci, 2006: 1528).
- ↑ Male, 2007: 319.
- ↑ Merino, Creagh y Martín, 2003: 129; Male, 2007: 319.
- ↑ The Body (2009), «Infección aguda del VIH» (Hoja 103E). Consultada el 14 de agosto de 2010.
- ↑ Daudel y Montagnier, 2002: 29-30.
- ↑ Dybull, Connors y Fauci, 2006: 1934.
- ↑ The Body (2009), «Infección aguda del VIH» (Hoja 103E). Consultada el 14 de agosto de 2010.
- ↑ "Clínica de la Infección por VIH-Sida", del Programa Municipal de Sida de Rosario (Argentina). Consultado el 10 de noviembre de 2010.
- ↑ Pérez, Cayón y Esquitino, 2003:86.
- ↑ Vélez, 2005: 5.
- ↑ Onusida, 1999.
- ↑ a b Sharp, P. M., Shaw, G. M. & Hahn, B. H. (2005). «Simian immunodeficiency virus infection of chimpanzees». J Virol 79: pp. 3891-902.
- ↑ Bailes et al., 2003.
- ↑ Levy, 2007: 22.
- ↑ Santiago, M. L., Range, F., Keele, B. F., Li, Y., Bailes, E., Bibollet-Ruche, F., Fruteau, C., Noe, R., Peeters, M., Brookfield, J. F., Shaw, G. M., Sharp, P. M. & Hahn, B. H. (2005). «Simian immunodeficiency virus infection in free-ranging sooty mangabeys (Cercocebus atys atys) from the Tai Forest, Cote d'Ivoire: implications for the origin of epidemic human immunodeficiency virus type 2». J Virol 79: pp. 12515-27.
- ↑ {{cita publicación|autor= Ling, B., Apetrei, C., Pandrea, I., Veazey, R. S., Lackner, A. A., Gormus, B. & Marx, P. A.|título= Classic AIDS in a sooty mangabey after an 18-year natural infection|revista= J Virol|año= 2004|páginas=
- ↑ Onusida (s/f): "Quiénes somos", en el portal de Onusida en español, consultado el 17 de noviembre de 2010.
- ↑ Inegi, 2007.
- ↑ Onusida, 2010: 32.
- ↑ Onusida, 2008: 39-47.
- ↑ Gómez C. E. et al. The HIV/ADS Vaccine Candidate MVA-B Administered as a Single Immunogen in Humans Triggers Robust, Polyfunctional, and Selective Effector Memory T Cell Responses to HIV-1 Antigens. Journal of Virology. DOI: 10.1128/jvi.05165-11
- ↑ García F. et al. Safety and immunogenicity of a modified pox vector-based HIV/AIDS vaccine candidate expressing ENV, Gag, Pol and Nef proteins of HIV-1 subtype B (MVA-B) in healthy HIV-uninfected volunteers: a phase I clinical trial. Vaccine. DOI: 10.1016/j.vaccine.2011.08.098
Bibliografía
- BAILES, E., GAO, F., BIBOLLET-RUCHE, F., COURGNAUD, V., PETERS, M., MARX, P. A., HAHN, B. H. y SHARP, P. M. (2003). «Hybrid origin of SIV in chimpanzees». Science 300: pp. 1713.
- BRAVO, Luis (2005), Manual de farmacoterapia. Madrid: Elsevier.
- DAUDEL, Raymond y Luc MONTAGNIER (2002), El SIDA. Ciudad de México: Siglo XXI.
- DYBULL, Mark; Mark CONNORS y Anthony FAUCI (2006), «Inmunología de la infección por VIH» en Mandell, Benett y Dolin (eds., 2006). pp. 1527-1546.
- GARCÍA F. et al. (2001), Safety and immunogenicity of a modified pox vector-based HIV/AIDS vaccine candidate expressing ENV, Gag, Pol and Nef proteins of HIV-1 subtype B (MVA-B) in healthy HIV-uninfected volunteers: a phase I clinical trial. Vaccine.
- GARRIDO, Armando; Rosa OLMO y Carlos Castel Aznar (2001), Bioquímica metabólica. Madrid: Tebar.
- GÓMEZ C. E. et al. (2011), The HIV/ADS Vaccine Candidate MVA-B Administered as a Single Immunogen in Humans Triggers Robust, Polyfunctional, and Selective Effector Memory T Cell Responses to HIV-1 Antigens. Journal of Virology.
- LEVY, Jay A. (2007), HIV and the pathogenesis of AIDS. Washington, D. C.: Wiley-Blackwell.
- INEGI (2007), "Estadísticas a propósito del Día Mundial de la Lucha contra el Sida. Datos nacionales", en el portal en internet del Inegi, consultado el 17 de noviembre de 2010.</ref>
- MALE, David (2007), Inmunología. Madrid: Elsevier España.
- MANDELL, Gerald; John BENETT y Raphael DOLIN (eds. 2006), Enfermedades infecciosas. Madrid: Elsevier España.
- MERINO, Dolores; Ricardo CREAGH y Ignacio MARTÍN (2003), «Infección aguda por el VIH» en Pachón, Pujol y Rivero (eds., 2003). pp. 129-136.
- ONUSIDA (1999), Enfermedades oportunistas relacionadas con el VIH. Consultada el 11 de noviembre de 2010.
- ----------- (2008), Informe sobre la epidemia mundial de sida. Consultada el 17 de noviembre de 2010.
- PASCUAL, Álvaro y CORRAL José Luis (2003), «El virus de inmunodeficiencia humana: inmunopatogenia» en Pachón, Pujol y Rivero (eds., 2003). pp. 29-36.
- PACHÓN, Jerónimo; Emilio PUJOL y Antonio RIVERO (eds., 2003). La infección por el VIH: guía práctica. Sociedad Andaluza de Enfermedades Infecciosas.
- PÉREZ, Salvador; Manuel CAYÓN y Begoña ESQUITINO (2003), «Historia natural y clasificación de la infeción por VIH-1» en Pachón, Pujol y Rivero (eds., 2003). pp. 85-94.
- VÉLEZ A., Hernán (2005), Manual de VIH/SIDA y otras infecciones de transmisión sexual. Bogotá: Corporación para Investigaciones Biológicas.
Enlaces externos
 Wikimedia Commons alberga contenido multimedia sobre virus de inmunodeficiencia humana. Commons
Wikimedia Commons alberga contenido multimedia sobre virus de inmunodeficiencia humana. Commons- En MedlinePlus puede encontrar más información sobre de la inmunodeficiencia humana Virus de la inmunodeficiencia humana
- Portal de Onusida en español
- Información sobre la prueba de VIH en América Latina





Wikimedia foundation. 2010.