- Complejo mayor de histocompatibilidad
-
El complejo mayor de histocompatibilidad (CMH o MHC, acrónimo para el inglés major histocompatibility complex), o complejo principal de histocompatibilidad, es una familia de genes ubicados en el brazo corto del cromosoma 6 cuyos productos están implicados en la presentación de antígenos a los linfocitos T .
En humanos, los genes MHC conforman el denominado sistema HLA (por human leukocyte antigen), porque estas proteínas se descubrieron como antígenos en los leucocitos, que podían detectarse con anticuerpos. Los genes MHC son fundamentales en la defensa inmunológica del organismo frente a los patógenos, y por otro lado, constituyen la principal barrera al trasplante de órganos y de células madre.
La región del brazo corto del cromosoma 6 que contiene los genes del MHC posee la información de:
- ciertas glucoproteínas de la membrana plasmática involucradas en los mecanismos de presentación y procesamiento de antígenos a los linfocitos T: se agrupan en los genes de clase II (que codifican las proteínas MHC-II) y los genes de clase I (que codifican las proteínas MHC-I)
- así como citocinas y proteínas del sistema del complemento, importantes en la respuesta inmunológica, pero que no tienen nada que ver con los genes del MHC; estos genes se agrupan en la clase III.
Ambos tipos de moléculas participan en la respuesta inmunitaria, que permite la identificación de las moléculas propias y de las extrañas (invasoras), para eliminar estas últimas mediante diferentes mecanismos.
Contenido
La región clases
Localización genómica
El análisis comparativo de la organización de la región MHC entre especies muy alejadas ha revelado la presencia de reorganizaciones dentro de la región específicas de cada línea evolutiva y cambios en la complejidad de los genes. La estructura de la región MHC se conoce al menos en siete especies de mamíferos euterios (placentarios), dos de aves, cinco peces teleósteos y en los tiburones. Hay grandes diferencias en la organización de la región MHC entre los mamíferos euterios y los no mamíferos. En euterios, la región está ordenada a lo largo del cromosoma en las regiones clase I-II-III, es muy densa en genes y ocupa una zona extensa. En no mamíferos, la región MHC generalmente contiene menos genes y las regiones Clase I y II son adyacentes, a excepción de los teleósteos, donde las dos regiones no están ligadas. De las regiones MHC secuenciadas completamente, la menos compleja es la del pollo, que contiene sólo 19 genes en 92 kb.[1]
En humanos, los 3,6 Mbp (3.600.000 pares de bases) de la región MHC del cromosoma 6 contiene 140 genes flanqueados por los marcadores genéticos MOG y COL11A2.[2] La región MHC es la región más densa en genes y más polimórfica del genoma de los mamíferos, crítica para la inmunidad y para el éxito reproductivo.
La región MHC en los marsupiales Monodelphis doméstica (Didelphimorphia gris de cola corta) está flanqueada por los mismos marcadores, comprende 3.95 Mb y contiene 114 genes, 87 compartidos con los humanos.[1] La comparación entre la región MHC de humanos y marsupiales ha permitido analizar la evolución de este conjunto de genes, ya que los marsupiales se encuentran entre los euterios y los vertebrados no mamíferos, separados por 200 millones de años. Así, se ha identificado que los marsupiales presentan una región MHC similar a la de mamíferos en tamaño y complejidad, pero también presenta características de organización similares a la región MHC de los no mamíferos, lo que revela una probable organización ancestral de esta región.
La región MHC está dividida en 3 subgrupos de genes:
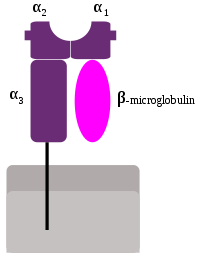 Estructura de una molécula MHC de Clase-I.
Estructura de una molécula MHC de Clase-I.
MHC Clase-I
En euterios, la región Clase-I contiene un conjunto de genes cuya presencia y orden está conservada entre especies. Estas moléculas se expresan en todas las células humanas, excepto en los glóbulos rojos, las células germinales, las células de los embriones pre-implantación y el sincitiotrofoblasto (tejido embrionario, no presente en la vida extrauterina: más detalles...).[3] Algunas células, como las neuronas, los monocitos y los hepatocitos, presentan niveles bajos de moléculas MHC-I (menos de 103 por célula: ver datos).[4]
Los genes MHC de Clase-I (MHC-I) codifican glucoproteínas, con estructura de inmunoglobulina: presentan una cadena pesada tipo α que se subdivide en tres regiones: α1, α2 y α3. Estas tres regiones están expuestas al espacio extracelular y están unidas a la membrana de la célula mediante una región transmembrana. La cadena α está siempre asociada a una molécula de microglobulina β2, que está codificada por una región independiente en el cromosoma 15.
La principal función de los productos génicos de la Clase-I es la presentación de péptidos antigénicos intracelulares a los linfocitos T citotóxicos (CD8+). El péptido antigénico se aloja en una hendidura que se forma entre las regiones α1 y α2 de la cadena pesada.
En humanos, existen muchos isotipos (genes diferentes) de las moléculas de Clase-I, que pueden agruparse en:
- "clásicas", cuya función consiste en la presentación de antígenos a los linfocitos T8: dentro de este grupo tenemos HLA-A, HLA-B y HLA-C.
- "no clásicas" (también llamadas MHC clase IB), con funciones especializadas: no presentan antígenos a los linfocitos T, sino que se unen a receptores inhibidores de las células NK; dentro de este grupo se encuentran HLA-E, HLA-F, HLA-G.
 Estructura de una molécula MHC de Clase-II.
Estructura de una molécula MHC de Clase-II.MHC Clase-II
Estos genes codifican glicoproteinas con estructura de inmunoglobulina, pero en este caso el complejo funcional está formado por dos cadenas, una α y una β (cada una de ellas con dos dominios, α1 y α2, β1 y β2). Cada una de las cadenas está unida a la membrana por una región transmembrana, y ambas cadenas están enfrentadas, con los dominios 1 y 2 contiguos, en el exterior celular.[5]
Estas moléculas se expresan sobre todo en las células presentadoras de antígeno (dendríticas y fagocíticas, así como los linfocitos B), donde presentan péptidos antigénicos extracelulares procesados a los linfocitos T cooperadores (CD4+). El péptido antigénico se aloja en una hendidura formada por los dominios α1 y β1.
Las moléculas MHC-II en humanos presentan 5-6 isotipos, y pueden agruparse en:
- "clásicas", que presentan péptidos a los linfocitos T4; dentro de este grupo tenemos HLA-DP, HLA-DQ, HLA-DR;
- "no clásicas", accesorias, con funciones intracelulares (no están expuestas en la membrana celular, sino en membranas internas, de los lisosomas); normalmente, cargan los péptidos antigénicos sobre las moléculas MHC-II clásicas; en este grupo se incluyen HLA-DM y HLA-DO.
Además de las moléculas MHC-II, en la región Clase-II se encuentran genes que codifican moléculas procesadoras de antígenos, como TAP (por transporter associated with antigen processing) y Tapasin.
MHC Clase-III
Esta clase contiene genes que codifican varias proteinas secretadas que desempeñan funciones inmunitarias: componentes del sistema del complemento (como C2, C4 y factor B) y moléculas relacionadas con la inflamación (citoquinas como TNF-α, LTA, LTB) o proteínas de choque térmico (hsp).
La Clase-III tiene una función completamente diferente que las clases- I y II, pero se sitúa entre los otros dos en el brazo corto del cromosoma 6 humano, por lo que son frecuentemente descritos en conjunto.
Polimorfismo de los genes MHC-I y II
 Expresión codominante de los genes HLA / MHC.
Expresión codominante de los genes HLA / MHC.Los genes MHC se expresan de forma codominante. Esto quiere decir que los alelos (variantes) heredados de ambos progenitores se expresan de forma equivalente:
- Como existen tres genes Clase-I, denominados en humanos HLA-A, HLA-B y HLA-C, y cada persona hereda un juego de cada progenitor, cualquier célula de un individuo podrá expresar 6 tipos diferentes de moléculas MHC-I.
- En el locus de Clase-II, cada individuo hereda un par de genes HLA-DP (DPA1 y DPA2, que codifican las cadenas α y β), un par de genes HLA-DQ (DQA1 y DQA2, para las cadenas α y β), un gen HLA-DRα (DRA1) y uno o dos genes HLA-DRβ (DRB1 y DRB3, -4 o -5). Así, un individuo heterocigoto puede heredar 6 u 8 alelos de Clase-II, tres o cuatro de cada progenitor.
El juego de alelos presente en cada cromosoma se denomina haplotipo MHC. En humanos, cada alelo HLA recibe un número. Por ejemplo, para un individuo dado, su haplotipo puede ser HLA-A2, HLA-B5, HLA-DR3, etc... Cada individuo heterocigoto tendrá dos haplotipos MHC, uno en cada cromosoma (uno de origen paterno y otro de origen materno).
Los genes MHC son enormemente polimórficos, lo que significa que existen muchos alelos diferentes en los diferentes individuos de la población. El polimorfismo es tan grande que en una población mixta (no endogámica) no existen dos individuos que tengan exactamente el mismo juego de genes y moléculas MHC, excepto los gemelos idénticos.
Las regiones polimórficas de cada alelo se encuentran en la zona de contacto con el péptido que va a presentar al linfocito. Por esta razón, la zona de contacto de cada alelo de molécula MHC es muy variable, ya que los residuos polimórficos del MHC forman hendiduras específicas en las que las que sólo pueden introducirse cierto tipo de residuo del péptido, lo cual impone un modo de unión muy preciso entre el péptido y la molécula MHC. Esto implica que cada variante de molécula MHC podrá unir específicamente sólo aquellos péptidos que encajen adecuadamente en la hendidura de la molécula MHC, que es variable para cada alelo. De esta manera, las moléculas de MHC tienen una especificidad amplia para la unión de péptidos, puesto que cada molécula de MHC puede unir muchos, pero no todos los tipos de péptidos posibles. Esta es una característica esencial de las moléculas MHC: en un individuo concreto, bastan unas pocas moléculas diferentes para poder presentar una vasta variedad de péptidos.
Por otro lado, dentro de una población, la existencia de múltiples alelos asegura que siempre habrá algún individuo que posea una molécula de MHC capaz de cargar el péptido adecuado para reconocer un microbio concreto.La evolución del polimorfismo de MHC asegura que una población será capaz de defenderse frente a la enorme diversidad de microbios existentes, y que no sucumbirá ante la presencia de un nuevo patógeno o un patógeno mutado, porque al menos algunos individuos serán capaces de desarrollar una respuesta inmune adecuada para vencer al patógeno. Las variaciones en las secuencias de MHC (responsables del polimorfismo) resultan de la herencia de diferentes moléculas MHC, y no son inducidas por recombinación, como ocurre con los receptores de los antígenos.
Funciones de las moléculas MHC-I y II
Ambos tipos de moléculas presentan péptidos antigénicos a los linfocitos T, responsables de la respuesta inmune específica para eliminar el patógeno responsable de la producción de dichos antígenos. Sin embargo, las moléculas MHC de Clase-I y II corresponden a dos vías diferentes de procesamiento de los antígenos, y se asocian con dos sistemas diferentes de defensa inmunitaria:[5]
Tabla 1. Características de las vías de procesamiento de antígenos Característica Vía MHC-II Vía MHC-I Composición del complejo estable péptido-MHC Cadenas polimórficas α y β, péptido unido a ambas Cadena polimórfica α y microglobulina β2, péptido unido a cadena α Tipos de células presentadoras de antígenos (APC) Células dendríticas, fagocitos mononucleares, linfocitos B, algunas células endoteliales, epitelio del timo Casi todas las células nucleadas Linfocitos T capaces de responder Linfocitos T cooperadores (CD4+) Linfocitos T citotóxicos (CD8+) Origen de las proteínas antigénicas Proteínas presentes en endosomas o lisosomas (en su mayoría internalizadas del medio extracelular) Proteínas citosólicas (en su mayor parte sintetizadas por la célula; pueden entrar también del exterior mediante fagosomas) Enzimas responsables de la generación de péptidos Proteasas de los endosomas y lisosomas (como la catepsina) El proteasoma citosólico Sitio de carga del péptido sobre la molécula MHC Compartimento vesicular especializado Retículo endoplásmico Moléculas implicadas en el transporte de los péptidos y carga sobre las moléculas MHC Cadena invariante, DM TAP (transporter associated with antigen processing)
Los linfocitos T de un individuo concreto presentan una propiedad denominada restricción MHC: sólo pueden detectar un antígeno si éste viene presentado por una molécula MHC del mismo individuo. Esto se debe a que cada linfocito T tiene una especificidad dual: el receptor del linfocito T (denominado TCR por T cell receptor) reconoce algunos residuos del péptido y simultáneamente algunos residuos de la molécula MHC que lo presenta. Esta propiedad es muy importante en el trasplante de órganos, e implica que, durante su desarrollo, los linfocitos T deben "aprender" a reconocer las moléculas MHC propias del individuo, mediante el proceso complejo de maduración y selección que tiene lugar en el timo.Las moléculas MHC sólo pueden presentar péptidos, lo que implica que los linfocitos T, dado que sólo pueden reconocer un antígeno si viene asociado a una molécula MHC, sólo pueden reaccionar ante antígenos de origen proteico (procedentes de microbios) y no a otro tipo de compuestos químicos (ni lípidos, ni ácidos nucleicos, ni azúcares). Cada molécula MHC puede presentar un único péptido cada vez, dado que la hendidura de la molécula sólo tiene espacio para alojar un péptido. Sin embargo, una molécula MHC dada tiene una especificidad amplia, porque puede presentar muchos péptidos diferentes (aunque no todos).
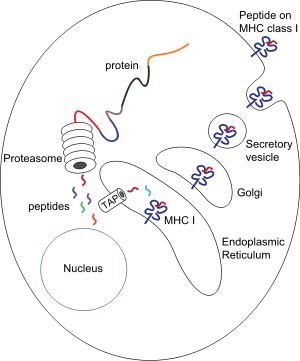 Procesamiento de péptidos asociados a moléculas MHC-I: proteínas presentes en el citosol son degradadas por el proteasoma, y los péptidos resultantes son internalizados por el canal TAP en el retículo endoplásmico, donde se asocian con las moléculas recién sintetizadas de MHC-I. Los complejos péptido-MHC-I pasan al aparato de Golgi, donde son glucosilados, y de ahí a vesículas secretoras, que se fusionan con la membrana celular, de forma que los complejos quedan expuestos hacia el exterior, permitiendo el contacto con los linfocitos T circulantes.
Procesamiento de péptidos asociados a moléculas MHC-I: proteínas presentes en el citosol son degradadas por el proteasoma, y los péptidos resultantes son internalizados por el canal TAP en el retículo endoplásmico, donde se asocian con las moléculas recién sintetizadas de MHC-I. Los complejos péptido-MHC-I pasan al aparato de Golgi, donde son glucosilados, y de ahí a vesículas secretoras, que se fusionan con la membrana celular, de forma que los complejos quedan expuestos hacia el exterior, permitiendo el contacto con los linfocitos T circulantes.Las moléculas MHC adquieren el péptido que presentan en el exterior de la membrana celular durante su propia biosíntesis, en el interior celular. Por tanto, los péptidos que presentan las moléculas MHC provienen de microbios que están en el interior celular, y ésta es la razón por la cual los linfocitos T, que sólo identifican péptidos cuando están asociados a moléculas MHC, sólo detectan microbios asociados a células y desencadenan una respuesta inmune contra microbios intracelulares.
Es importante resaltar que las moléculas MHC-I adquieren péptidos que provienen de proteínas citosólicas, mientras que las moléculas MHC-II adquieren péptidos de proteínas contenidas en vesículas intracelulares. Por ello, las moléculas MHC-I presentarán péptidos propios, péptidos virales (sintetizados por la propia célula) o péptidos procedentes de microbios ingeridos en fagosomas. Las moléculas MHC-II, por su parte, presentarán péptidos procedentes de microbios ingeridos en vesículas (este tipo de moléculas sólo se expresan en células con capacidad fagocítica). Las moléculas MHC sólo se expresan de forma estable en la membrana celular si tienen un péptido cargado: la presencia del péptido estabiliza la estructura de las moléculas MHC, las moléculas "vacías" se degradan en el interior celular. Las moléculas MHC cargadas con un péptido pueden permanecer en la membrana durante días, el tiempo suficiente para asegurar que un linfocito T adecuado reconozca el complejo e inicie la respuesta inmunitaria.
En cada individuo, las moléculas MHC pueden presentar tanto péptidos extraños (procedentes de patógenos) como péptidos procedentes de las proteínas propias del individuo. Esto implica que, en un momento dado, sólo una pequeña proporción de las moléculas MHC de una célula presentará un péptido extraño: la mayor parte de los péptidos que presente serán propios, dado que son más abundantes. Sin embargo, los linfocitos T son capaces de detectar un péptido presentado por sólo el 0,1%-1% de las moléculas MHC para iniciar una respuesta inmune.
Los péptidos propios, por otro lado, no pueden iniciar una respuesta inmune (excepto en los casos de las enfermedades autoinmunes), porque los linfocitos T específicos para los antígenos propios son destruidos o inactivados en el timo. Sin embargo, la presencia de péptidos propios asociados a las moléculas MHC es esencial para la función supervisora de los linfocitos T: estas células están constantemente patrullando el organismo, verificando la presencia de péptidos propios asociados a las moléculas MHC y desencadenando una respuesta inmune en los casos raros en los que detectan un péptido extraño.
Las moléculas MHC en el rechazo de trasplantes
Las moléculas MHC se identificaron y nombraron precisamente por su papel en el rechazo de trasplantes entre ratones de diferentes cepas endogámicas. En humanos, las moléculas MHC son los antígenos de los leucocitos (HLA). Llevó más de 20 años comprender la función fisiológica de las moléculas MHC en la presentación de péptidos a los linfocitos T.[6]
Como se ha indicado antes, cada célula humana expresa 6 alelos MHC de clase-I (un alelo HLA-A, -B y -C de cada progenitor) y 6-8 alelos MHC de clase-2 (uno HLA-DP y -DQ, y uno o dos de HLA-DR de cada progenitor, y algunas combinaciones de éstos). El polimorfismo de los genes MHC es muy elevado: se estima que en la población hay al menos 350 alelos de los genes HLA-A, 620 alelos de HLA-B, 400 alelos de DR y y 90 alelos de DQ. Como estos alelos pueden heredarse y expresarse en muchas combinaciones diferentes, cada individuo expresará probablemente algunas moléculas que serán diferentes de las moléculas de otro individuo, excepto en el caso de los gemelos idénticos. Todas las moléculas MHC pueden ser dianas del rechazo de transplantes, aunque HLA-C y HLA-DP tienen un bajo polimorfismo, y probablemente tengan una importancia menor en los rechazos.
En el caso de un trasplante (de órganos o de células madre), las moléculas HLA funcionan como antígenos: pueden desencadenar una respuesta inmunitaria en el receptor, provocando el rechazo del trasplante. El reconocimiento de los antígenos MHC en células de otro individuo es una de las respuestas inmunes más intensas que se conocen. La razón por la que los individuos reaccionan contra las moléculas MHC de otro individuo se comprende bastante bien.
Durante el proceso de maduración de los linfocitos T, éstos son seleccionados en función de la capacidad de su TCR de reconocer débilmente complejos "péptido propio:MHC propio". Por ello, en principio, los linfocitos T no deberían reaccionar frente a un complejo "péptido extraño:MHC extraño", que es lo que aparecerá en las células transplantadas. Sin embargo, parece que lo que ocurre es un tipo de reacción cruzada: los linfocitos T del individuo receptor pueden equivocarse, porque la molécula MHC del donante es similar a la propia en la zona de unión al TCR (la zona variable del MHC se encuentra en la zona de unión al péptido que presentan). Por esta razón, los linfocitos del individuo receptor interpretan el complejo presente en las células del órgano trasplantado como "péptido extraño:MHC propio" y desencadenan una respuesta inmune contra el órgano "invasor", porque lo perciben de la misma manera que un tejido propio infectado o tumoral, pero con un número mucho más elevado de complejos capaces de iniciar una respuesta. El reconocimiento de la molécula de MHC extraña como propia por los linfocitos T se denomina aloreconocimiento.
Pueden producirse dos tipos de rechazo de trasplantes mediado por las moléculas MHC (HLA):
- rechazo hiperagudo: se produce cuando el individuo receptor presenta anticuerpos anti-HLA preformados, antes del trasplante; esto puede deberse a la realización previa de transfusiones de sangre (que incluye linfocitos del donante, con moléculas HLA), a la generación de anti-HLA durante el embarazo (contra los HLA del padre presentes en el feto) y por la realización de un trasplante previo;
- rechazo humoral agudo y disfunción crónica del órgano trasplantado: se debe a la formación de anticuerpos anti-HLA en el receptor, contra las moléculas HLA presentes en las células endoteliales del trasplante.
En ambos casos, se produce una reacción inmune contra el órgano trasplantado, que puede generar lesiones en el mismo, lo que conlleva la pérdida de función, inmediata en el primer caso y progresiva en el segundo.
Por esta razón, es fundamental realizar una prueba de reacción cruzada entre células del donante y suero del receptor, para detectar la presencia de anticuerpos anti-HLA preformados en el receptor contra las moléculas HLA del donante, y evitar el rechazo hiperagudo. Normalmente, se verifica la compatibilidad de las moléculas HLA-A, -B y -DR: a medida que aumenta el número de incompatibilidades, la supervivencia a 5 años del transplante disminuye. La compatibilidad total sólo existe entre gemelos idénticos, pero en la actualidad existen bases de datos de donantes a nivel mundial que permiten optimizar la compatibilidad HLA entre un donante potencial y un receptor.
Referencias
- ↑ a b Belov, Katherine; Janine E. Deakin, Anthony T. Papenfuss, et. al (March 2006). «Reconstructing an Ancestral Mammalian Immune Supercomplex from a Marsupial Major Histocompatibility Complex». PLoS Biol 4(3) (e46). doi:.
- ↑ MHC Sequencing Consortium (1999). «Complete sequence and gene map of a human major histocompatibility complex». Nature 401: pp. 921–923.
- ↑ Comiskey, M.; Warner C.M, Schust D.J. (2000). Landes Bioscience. ed. MHC Molecules of the Preimplantation Embryo and Trophoblast. Madame Curie Bioscience Database [Internet].. NBK6277.
- ↑ Berke, G.; Clark W.R. (2007). «Ch.1 Basic Immunology: A Primer». Killer lymphocytes (2nd edición). Springers. ISBN 9781402065637.
- ↑ a b Abbas, A.B.; Lichtman A.H. (2009). «Ch.3 Antigen capture and presentation to lymphocytes». Basic Immunology. Functions and disorders of the immune system (3rd edición). Saunders (Elsevier). ISBN 978-1-4160-4688-2.
- ↑ Abbas, A.B.; Lichtman A.H. (2009). «Ch.10 Immune responses against tumors and transplants». Basic Immunology. Functions and disorders of the immune system (3rd edición). Saunders (Elsevier). ISBN 978-1-4160-4688-2.
Véase también
Categorías:- Genética
- Glicoproteínas
- Antígenos




Wikimedia foundation. 2010.