- Liliaceae
-
 Liliaceae
Liliaceae
Lilium henryi.Clasificación científica Reino: Plantae División: Angiospermae Clase: Monocotyledoneae Orden: Liliales Familia: Liliaceae Juss.[1] géneros Ver Anexo:Géneros y especies tradicionalmente incluidos en Liliaceae para un listado de algunos de los géneros y especies que se incluían en una definición amplia de Liliaceae. La circunscripción actual de Liliaceae es la siguiente:
- Calochortus (incl. Cyclobothra).
- Cardiocrinum
- Clintonia
- Erythronium
- Fritillaria (incl. Orithyia, Petilium, Rhinopetalum, Korolkowia, Theresia)
- Gagea
- Lilium
- Lloydia (incl. Giraldiella).
- Medeola
- Nomocharis
- Notholirion
- Prosartes(sin. Disporum)
- Scoliopus
- Streptopus
- Tricyrtis (incl. Brachycyrtis, Compsoa')
- Tulipa (incl. Amana)
Las liliáceas (nombre científico Liliaceae Juss.) son una familia de plantas monocotiledóneas perennes, herbáceas, con frecuencia bulbosas, que pueden ser reconocidas por sus flores bastante grandes con un perigonio formado por 6 tépalos libres, frecuentemente coloreados y con manchas, 6 estambres extrorsos y un ovario súpero, tricarpelar y trilocular. Se hallan ampliamente distribuidas por todo el mundo, principalmente en regiones templadas del hemisferio norte.[2] La familia fue reconocida por sistemas de clasificación modernos como el sistema de clasificación APG II del 2003[3] y el APWeb (2001 en adelante[2] ), pero la circunscripción moderna de la familia es mucho más estricta que la tradicionalmente aceptada, para que se mantenga monofilética en base a los análisis moleculares de ADN recientes. Como aquí circunscripta, consta de cerca de 16 géneros y 600 especies.
Contenido
Antecedentes históricos
La familia Liliaceae fue concebida por Antoine Laurent de Jussieu en 1789 y su definición era muy amplia y artificial: todas las especies de plantas con 6 tépalos y gineceo de ovario súpero eran incluidas en esta familia. En un momento llegó a abarcar cerca de 300 géneros y 4.500 especies y se incluían dentro del gran orden Liliales (Cronquist 1981, Thorne 1992)[4] [5] como las monocotiledóneas petaloideas, un "grupo" caracterizado por flores con tépalos vistosos y sin almidón en el endosperma. Cronquist (1981)[4] ubicó a la mayoría de las monocotiledóneas petaloideas con flores de 6 estambres en un muy amplio (y ahora sabemos que ampliamente polifilético) Liliaceae. Mientras que, en otros tratamientos, de han dividido las monocotiledóneas petaloideas con 6 estambres en Liliaceae, incluyendo especies con un ovario súpero, y Amaryllidaceae, que incluye a las especies con ovario ínfero (Lawrence 1951).[6] No obstante, con el tiempo se llegó a reconocer que, definida de esa forma, la familia Liliaceae incluía un vasto y heterogéneo repertorio de géneros que no estaban relacionados filogenéticamente. Existieron varias propuestas para separar grupos pequeños de géneros en familias más homogéneas, pero ninguna de ellas fue ampliamente aceptada. En la década de 1980, en el contexto de una revisión general de la clasificación de las angiospermas, las liliáceas fueron sometidas a un escrutinio más intenso. Hacia fines de esa década, los Jardines Botánicos de Kew, el Museo Británico de Ciencias Naturales y los Jardines Botánicos de Edimburgo formaron un comité para examinar la posibilidad de separar a las liliáceas en subgrupos más homogéneos, al menos para la organización de sus herbarios. El comité recomendó que se utilicen 24 nuevas familias en lugar de la vieja y ampliamente definida Liliaceae.[7] En las últimas dos décadas los estudios de ADN y los datos morfológicos (particularmente aquellos relacionados con la morfología reproductiva) sumados a los análisis cladísticos, han permitido concluir que las "monocotiledóneas petaloideas" en realidad no pertenecen a una misma familia botánica sino que se distribuyen en dos órdenes diferentes: Asparagales y Liliales. La monofilia de estos órdenes está sustentada por análisis cladísticos basados en morfología, ADNr 18S, y muchas otras secuencias de ADN.[8] [9] [10] [11] [12] [13] [14] [15] [16] [17] [18] [19] [20] [21] [22] [17] [23] [24]
Tales estudios, sumados a los análisis realizados dentro de cada uno de los órdenes han permitido reagrupar a los géneros previamente incluidos dentro de Liliaceae en una gran cantidad de familias, como se muestra a continuación a través de los cladogramas para los Asparagales y Liliales (según Judd et al. 2007, modificado de Soltis et al. 2005):[25]
Asparagales Hypoxidaceae (tradicionalmente ubicado en Amaryllidaceae)
Xanthorrhoeaceae sensu lato Hemerocallidaceae (tradicionalmente ubicado en Liliaceae)
Xanthorrhoeaceae (tradicionalmente ubicado en Liliaceae)
Asphodelaceae (tradicionalmente ubicado en Liliaceae)
Alliaceae sensu lato Agapanthaceae (tradicionalmente ubicado en Liliaceae)
Alliaceae (tradicionalmente ubicado en Liliaceae)
Laxmanniaceae (tradicionalmente ubicado en Liliaceae)
Asparagaceae (tradicionalmente ubicado en Liliaceae)
Ruscaceae (tradicionalmente ubicado en Liliaceae)
Themidaceae (tradicionalmente ubicado en Liliaceae)
Hyacinthaceae (tradicionalmente ubicado en Liliaceae)
Agavaceae (tradicionalmente ubicado en Amaryllidaceae o Liliaceae)
Liliales Alstroemeriaceae (tradicionalmente ubicado en Liliaceae)
Colchicaceae (tradicionalmente ubicado en Liliaceae)
Melanthiaceae (tradicionalmente ubicado en Liliaceae)
Smilacaceae (tradicionalmente ubicado en Liliaceae)
Liliaceae en sentido estricto
Ambos cladogramas demuestran que la definición clásica de Liliaceae era incorrecta, sumamente artificial y no reflejaba las verdaderas relaciones filogenéticas entre las especies que la componían. Por esta razón, y pese al uso extendido de la circunscripción amplia de las liliáceas, los botánicos de todo el mundo fueron adoptando la definición más estricta de esta familia y la segregación de los géneros que la componían en dos órdenes y numerosas familias.
Descripción
 Fritillaria meleagris, sección de la flor.
Fritillaria meleagris, sección de la flor.
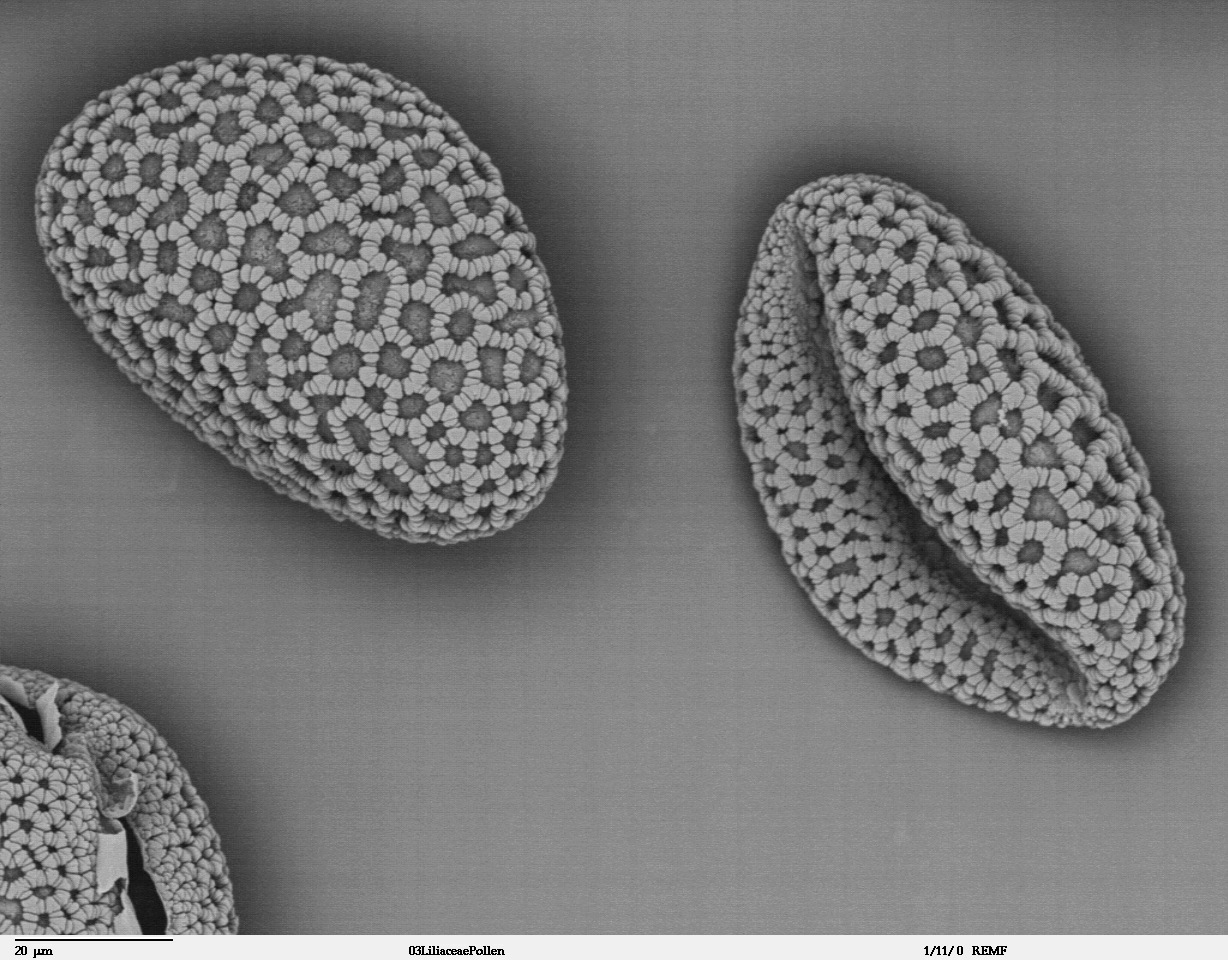
 Granos de polen de Lilium auratum.
Granos de polen de Lilium auratum. Flor de Calochortus nuttallii, obsérvense las diferencias morfológicas existentes entre los dos verticilos de tépalos.
Flor de Calochortus nuttallii, obsérvense las diferencias morfológicas existentes entre los dos verticilos de tépalos.Hierbas perennes con bulbos o rizomas, con pelos simples. Las raíces son típicamente contráctiles.
Las hojas son alternas y espiraladas (e incluso verticiladas, como en Lilium y Fritillaria) y se disponen a lo largo del tallo o en una roseta basal. Son simples, enteras, con venación paralela (aunque en Prosartes y Tricyrtis con venación claramente reticulada entre las venas primarias), muchas veces son envainadoras en la base. Raramente las hojas son pecioladas. No presentan estípulas.
La inflorescencia es usualmente determinada, a veces reducida a una única flor, y terminal. Cuando es multiflora las flores se disponen en un racimo o raramente en una umbela. Las flores son hermafroditas, actinomorfas o ligeramente zigomorfas, en general grandes y vistosas, pediceladas, pueden o no presentar brácteas. El perigonio está formado por 6 tépalos dispuestos en dos verticilos trímeros, se hallan separados entre sí y libres de las demás piezas florales, imbricados, son petaloideos y usualmente presentan manchas, puntos o líneas de otros colores o tonos. El perianto puede ser homoclamídeo (o sea, todos los tépalos son iguales entre sí, como en Fritillaria) o diclamídeo (los dos verticilos de tépalos presentan diferencias morfológicas, como en Calochortus). El néctar se produce en nectarios en la base de los tépalos.
El androceo presenta 6 estambres dispuestos en 2 verticilos también trímeros, los filamentos se hallan separados entre sí y libres de las demás piezas florales. Característicamente, el androceo es diplostémono (es decir que el verticilo externo de estambres es opuesto a los tépalos externos y el ciclo interno es opuesto a los tépalos internos). Las anteras están unidas al filamento en forma peltada o pseudo-basifijas (la punta del filamento rodeada pero no adherida al tejido conectivo), y de dehiscencia longitudinal. El polen es en general monosulcado.
El gineceo es de ovario súpero y está formado por 3 carpelos connados, es trilocular. Presenta un solo estilo y un estigma 3-lobado o bien, 3 estigmas más o menos elongados que se extienden a lo largo de la cara interna de las ramas del estilo. Los óvulos son numerosos, con placentación axilar, usualmente con un tegumento y un megasporangio más o menos delgado. El saco embrionario (megagametofito o gametofito femenino) es variable según el género considerado. Puede ser monospórico (tipo Polygonum) o tetraspórico (tipo Fritillaria). [a]La plantilla {{Note label}} está obsoleta, véase el nuevo sistema de referencias.
El fruto es una cápsula loculicida o septicida, ocasionalmente una baya. Las semillas son planas y con forma de disco o globosas, el tegumento no es negro, rasgo que las diferencia de otras familias, como por ejemplo Alliaceae. El endosperma es aceitoso (con aleurona y aceites) pero sin almidón, sus células pueden ser triploides o pentaploides, dependiendo del saco embrionario considerado.
Desde un punto de vista fitoquímico, la familia presenta saponinas esteroideas pero no exhibe cristales de rafidio ni ácido chelidónico ni compuestos azufrados derivados de la cisteína (o sea, no tienen el olor aliáceo característico de los ajos y las cebollas).
Los números cromosómicos básicos son variables de acuerdo al género considerado.
Ver un tratamiento reciente de la familia en Tamura (1998b).[26]
Ecología
Las liliáceas están ampliamente distribuidas, principalmente en regiones templadas del hemisferio norte. En general son plantas de praderas llanas, prados de montaña y otras comunidades abiertas. Poseen su centro de diversidad en el sudoeste de Asia a China.
Suelen florecer en la primavera. Las vistosas flores de esta familia son polinizadas por insectos, especialmente abejas, avispas, mariposas y polillas. El néctar o polen que producen las flores de las liliáceas en gran cantidad son empleados como recompensa de la polinización.
Las semillas son dispersadas tanto por el viento como por el agua, unas pocas especies tienen estructuras de tipo arilo y son dispersadas por hormigas.
Filogenia
 Nomocharis aperta en su hábitat natural, Napa Hai, Yunnan, China.
Nomocharis aperta en su hábitat natural, Napa Hai, Yunnan, China. Gagea lutea.
Gagea lutea. Clintonia borealis, fruto baya.
Clintonia borealis, fruto baya.Liliaceae, en su actual definición estricta, es claramente monofilética (Chase et al. 1995a,[8] b[9] ), si bien es difícil de diagnosticar morfológicamente (Tamura 1998b [26] ). Hayashi y Kawano (2000[27] ) secuenciaron las regiones rbcL y matK del ADN cloroplastídico de Lilium y géneros relacionados y encontraron resultados coherentes con la circunscripción de Liliaceae sensu stricto propuesta por Tamura (1998,[26] quien separó la subfamilia Calochortoideae en su propia familia Calochortaceae). Patterson y Givnish (2002[28] ) secuenciaron los genes rbcL y ndhF y encontraron un fuerte sostén para la monofilia de Liliaceae, la publicación también describe una filogenia de los géneros y provee estimaciones de los tiempos de divergencia.
Relacionadas con esta familia están Philesiaceae (2 géneros monoespecíficos, del sur de Sudamérica), Smilacaceae (monogenérica, 315 especies, casi cosmopolita), y Rhipogonaceae (1 género con 6 especies, del este de Australia), como avalado en Chase et al. 1995a,[8] 2006,[11] Conran 1998,[29] Fay et al. 2006,[30] Rudall et al. 2000a.[31] No se conocen caracteres fuera de los del ADN que unan a todas estas familias. Smilax es en general hermano de Liliaceae (pero nunca con más de 80 % de apoyo). Fay et al. (2006:[30] bajo apoyo), Givnish et al. (2006:[32] alto apoyo), y Chase et al. (2006[11] ) encontraron a Philesiaceae y Rhipogonaceae como taxones hermanos, y a Smilacaceae como hermano de Liliaceae.
Calochortus, Prosartes, Scoliopus, Streptopus, y Tricyrtis forman un clado (la subfamilia Calochortoideae), y son hierbas con rizomas rastreros ("creeping"), estilos divididos en el ápice, y el desarrollo del megagametofito del tipo Polygonum (es decir, el megagametofito desarrollándose de una megaspora simple, con endosperma triploide). En otras clasificaciones Calochortus ha sido ubicada en su propia familia, mientras que los demás géneros de Calochortoideae fueron ubicados en un heterogéneo Uvulariaceae (Dahlgren et al. 1985[33] ) o en un Calochortaceae expandido (Tamura 1998a[34] ). Sin embargo, estos miembros de Liliaceae no están cercanamente relacionados con los morfológicamente similares Uvularia y Disporum (Shinwari et al. 1994[35] ), y el último es aquí ubicado en Colchicaceae.
El resto de los géneros de Liliaceae constituyen los Lilioideae, un gran clado caracterizado por bulbos y raíces contráctiles, y un megagemetofito desarrollándose de 4 megasporas (de tipo Fritillaria).
La monofilia de cada uno de estos dos subclados de Liliaceae está sostenida por caracteres de secuencias de ADN (Chase et al. 1995a,[8] 2000,[10] Patterson y Givnish 2002[28] ).
Un tercer clado fue identificado a partir de las investigaciones filogenéticas de Clintonia, un género de distribución disyunta entre América del Norte y el este de Asia. Utilizando las 5 especies de Clintonia (Clintonia andrewsiana, Clintonia borealis, Clintonia umbellulata, Clintonia uniflora y Clintonia udensis), varios representantes de Liliaceae (tales como Cardiocrinum cordatum, Medeola virginiana, Scoliopus bigelovii y Scoliopus hallii) y los datos de las secuencias de los genes cloroplastídicos rbcL y matK, se llegó a la conclusión que Clintona es monofilético, que consiste en dos clados, uno de Asia oriental y el otro de Norteamérica, y que el género más estrechamente relacionado es Medeola. Ambos géneros, Clintonia y Medeola, fueron dispuestos en una subfamilia separada, Medeoloideae.[36]
Taxonomía
- Introducción teórica en Taxonomía
Las subfamilias de Liliaceae
- 1. Lilioideae Eaton (sinónimos: Erythroniaceae Martynov, Fritillariaceae R.A.Salisbury, Liriaceae Batsch, Medeolaceae Takhtajan, Tulipaceae Batsch). Incluye plantas frecuentemente bulbosas y con raíces contráctiles. Los tallos no son ramificados, las hojas tienen nerviación paralela. Las flores son grandes y el saco embrionario es del tipo Fritillaria(o sea, tetraspórico). El fruto es una cápsula septicida. Las semillas son frecuentemente aplanadas, con la exotesta en empalizada o lignificada. El número cromosómico básico es variable (n=9, 11 a 14). Los cromosomas, a su vez, presentan una longitud muy variable, que va desde los 2,2 µm hasta los 27 µm. Esta subfamilia comprende 11 géneros y alrededor de 535 especies que se distribuyen en regiones templadas y frías de América del Norte y el este de Asia. Los géneros de Lilioideae con mayor riqueza de especies son Tulipa (150 especies), Fritillaria (130), Lilium (110) y Gagea (90).
- 2. Calochortoideae Dumortier. Presentan hojas con nerviación paralela, flores grandes, sin estilo o con un estilo muy corto. Poseen un perianto diferenciado en cáliz y corola, con los tépalos pubescentes. El fruto es una cápsula con dehiscencia septicida y el saco embrionario es del tipo Polygonum.[37] [38] Los números cromosómicos varían de n=6 a n=13, con los cromosomas de 1,5 a 6,5 µm de longitud. Comprende 5 géneros, Calochortus, Prosartes, Scoliopus, Streptopus y Tricyrtis, y unas 100 especies distribuidas en el este de América del Norte y en el este de Asia. Varios taxónomos han sugerido que estos 5 géneros deben ser segregados de Liliaceae y disponerse en su propia familia, Calochortaceae.[39] [40] No obstante lo anterior, los sistemas modernos de clasificación (como APG o APG II o APWeb) no reconocen a esta última familia e incluyen a Calochortus y géneros afines dentro de las Liliáceas.
- 3. Medeoloideae . Incluye 2 géneros (Clintonia y Medeola) y unas 7 especies distribuidas en el este de Asia y en América del Norte. Presentan semillas con estrías y un número cromosómico básico de n=7.[36]
Listado de géneros
 Streptopus streptopoides var. japonicus.
Streptopus streptopoides var. japonicus.Como fue descrito en los antecedentes históricos, la definición de Liliaceae ha cambiado en el transcurso de los años. En el pasado se trató a la familia como un gran conjunto heterogéneo (Liliaceae sensu lato) que, más recientemente, fue dividido en numerosas familias segregadas. En la circunscripción moderna de Liliaceae se considera que la familia está compuesta por 16 géneros y 635 especies, los cuales se listan a continuación:[2] [41]
- Calochortus Pursh (incl.: Cyclobothra Sweet).
- Cardiocrinum (Endl.) Lindl.
- Clintonia Raf.
- Erythronium L.
- Fritillaria L. (incl.: Orithyia D.Don, Petilium Ludw., Rhinopetalum Fisch. ex D.Don, Korolkowia Regel, Theresia K.Koch)
- Gagea Salisb.
- Lilium L.
- Lloydia Salisb. ex Rchb. (incl.: Giraldiella Dammer).
- Medeola L.
- Nomocharis Franch.
- Notholirion Wall. ex Boiss.
- Prosartes D.Don (sin.: Disporum Salisb.)
- Scoliopus Torr.
- Streptopus Michx.
- Tricyrtis Wall. (incl.: Brachycyrtis Koidz., Compsoa D.Don)
- Tulipa L. (incl.: Amana Honda)
Los géneros más representados son Fritillaria (100 especies), Gagea (90 especies), Tulipa (80 especies), Lilium (80 especies), y Calochortus (65 especies).
Importancia económica
 Tricyrtis, detalle de la flor, las manchas de colores o tonos diferentes en los tépalos son bastante usuales en las Liliáceas.
Tricyrtis, detalle de la flor, las manchas de colores o tonos diferentes en los tépalos son bastante usuales en las Liliáceas. Los tulipanes (Tulipa sp.) son muy apreciados como flor de corte.
Los tulipanes (Tulipa sp.) son muy apreciados como flor de corte.- Véase también: Plantas bulbosas, Floricultura
Muchas especies de los géneros Tulipa ("tulipán"), Fritillaria ("ajedrezada"), Lilium ("lirio", "azucena"), Calochortus y Erythronium son cultivadas como plantas ornamentales en todo el mundo. No obstante, los tulipanes y las azucenas o lirios son las de mayor importancia económica dentro de la familia, como se describe a continuación.
Tulipán
El tulipán se cultiva con dos objetivos principales, la producción de flor cortada y la de bulbos secos. Estos últimos se destinan, a su vez, a satisfacer la demanda de bulbos para parques, jardines y uso hogareño y, por otro lado, para proveer los bulbos necesarios para la producción de flor cortada. El comercio internacional de flor cortada tiene un valor global aproximado de 11.000 millones de Euros, lo cual provee una magnitud de la importancia económica de esta actividad.
El principal país productor de bulbos de tulipán es Holanda, país que concentra el 87% del área mundial cultivada, la cual es de aproximadamente 12.000 hectáreas. Los bulbos de esta especie se producen significativamente en otros 14 países, encabezados por Japón, Francia y Polonia. La mayoría de estos países utiliza los bulbos obtenidos para su propia producción de flor cortada o para abastecer su mercado minorista de bulbos secos. Holanda, sin embargo, aparte de ser el principal productor internacional de bulbos, es la excepción a esta generalización. De hecho, produce aproximadamente 4.000 millones de bulbos anualmente, de los cuales el 53% se usan en el mercado de flor cortada y los restantes se utilizan en el mercado de bulbos secos. De los bulbos destinados al mercado de flor cortada, Holanda utiliza el 57% para satisfacer su mercado interno y el resto lo exporta a varios países, dentro y fuera de la Unión Europea.[42]
Lirios y azucenas
De modo similar que en el caso de los tulipanes, la mayor área de producción de las variedades de Lilium se concentra e Holanda (76% de área mundial). Otros 9 países, encabezados por Francia, Chile, Japón, Estados Unidos y Nueva Zelanda, producen azucenas a escala significativa. La mitad de los países productores utilizan los bulbos que producen en la industria de la floricultura y sólo una pequeña proporción se destina al mercado de ventas de bulbos secos. Países tales como Holanda, Francia, Chile, Nueva Zelanda y Australia utilizan los bulbos producidos para abastecer no sólo su mercado interno sino también para exportarlos. Holanda produce alrededor de 2.200 millones de bulbos de lirio anualmente, de los cuales el 96% se utiliza para su propia producción de flores y el resto es exportado a países de la Unión Europea principalmente.[42]
Véase también
Notas
a. ↑ La plantilla {{Note label}} está obsoleta, véase el nuevo sistema de referencias. Los sacos embrionarios pueden ser de diferentes tipos de acuerdo a cuántas megasporas degeneren después de la meiosis que sufre la célula madre de la megaspora. En el tipo Polygonum degeneran tres de las 4 megasporas y por eso el saco embrionario se dice monospórico (proviene de una sola espora). En el saco embrionario de tipo Fritillaria ninguna de las 4 megasporas degenera luego de la meiosis y todas ellas participan en la estructura del saco embrionario maduro, por eso se denomina "tetraspórico".[43]
Referencias citadas
- ↑ Jussieu, A. L., . (1789). Genera Plantarum, 48.
- ↑ a b c d Stevens, P. F. (2001 en adelante). «Liliaceae» (en inglés). Angiosperm Phylogeny Website. Versión 7, Mayo 2006. Consultado el 28 de abril de 2008.
- ↑ APG II (2003). «An Update of the Angiosperm Phylogeny Group Classification for the orders and families of flowering plants: APG II.» (pdf). Botanical Journal of the Linnean Society (141): pp. 399-436. http://www.blackwell-synergy.com/doi/pdf/10.1046/j.1095-8339.2003.t01-1-00158.x. Consultado el 12/01/2009.
- ↑ a b Cronquist, A. (1981). An integrated system of classification of flowering plants.. Nueva York: Columbia University Press.
- ↑ Thorne, R. F. (1992). «Classification and geography of the flowering plants.». Bot. Rev. (58): pp. 225-348.
- ↑ Lawrence, G. H. M. (1951). Taxonomy of vascular plants. (Macmillan edición). New York.
- ↑ Mathew, Brian (1989). Splitting the Liliaceae. The Plantsman 11 (2): 89.
- ↑ a b c d Chase, M. W.; Duvall, M. R., Hills, H. G., Conran, J. G., Cox, A. V., Eguiarte, L. E., Hartwell, J., Fay, M. F., Caddick, L. R., Cameron, K. M., y Hoot, S. (1995). «Molecular systematics of Lilianae.». En Rudall, P. J., Cribb, P. J., Cutler, D. F.. Monocotyledons: Systematics and evolution. (Royal Botanic Gardens edición). Kew. pp. 109-137.
- ↑ a b Chase, M. W.; Stevenson, D. W., Wilkin, P., y Rudall, P. J. (1995b). «Monocot systematics: A combined analysis.». En Rudall, P. J., Cribb, P. J., Cutler, D. F.. Monocotyledons: Systematics and evolution. (Royal Botanic Gardens edición). Kew. pp. 685-730.
- ↑ a b Chase, M. W.; Soltis, D. E., Soltis, P. S., Rudall, P. J., Fay, M. F., Hahn, W. H., Sullivan, S., Joseph, J., Molvray, M., Kores, P. J., Givnish, T. J., Sytsma, K. J., y Pires, J. C. (2000). «Higher-level systematics of the monocotyledons: An assessment of current knowledge and a new classification.». En Wilson, K. L. y Morrison, D. A.. Monocots: Systematics and evolution. (CSIRO Publ. edición). Collingwood, Australia. pp. 3-16.
- ↑ a b c Chase, M. W.; Fay, M. F.; Devey, D. S.; Maurin, O; Rønsted, N; Davies, T. J; Pillon, Y; Petersen, G; Seberg, O; Tamura, M. N.; Lange, Conny Bruun Asmussen (Faggruppe Botanik); Hilu, K; Borsch, T; Davis, J. I; Stevenson, D. W.; Pires, J. C.; Givnish, T. J.; Sytsma, K. J.; McPherson, M. A.; Graham, S. W.; Rai, H. S. (2006). «Multigene analyses of monocot relationships : a summary» (pdf). Aliso (22): pp. 63-75. ISSN: 00656275. http://www.ninaronsted.dk/page1/files/page1_3.pdf. Consultado el 25/02/2008.
- ↑ Davis, J. I.; Stevenson, D. W.; Petersen, G.; Seberg, O.; Campbell, L. M.; Freudenstein, J. V.; Goldman, D. H.; Hardy, C. R.; Michelangeli, F. A.; Simmons, M. P.; Specht, C. D.; Vergara-Silva, F.; Gandolfo, M. (2004). «A phylogeny of the monocots, as inferred from rbcL and atpA sequence variation, and a comparison of methods for calculating jacknife and bootstrap values.». Syst. Bot. (29): pp. 467-510. http://www.ingentaconnect.com/content/aspt/sb/2004/00000029/00000003/art00001. Consultado el 25/02/2008.
- ↑ Graham, S. W.; Zgurski, J. M., McPherson, M. A., Cherniawsky, D. M., Saarela, J. M., Horne, E. S. C., Smith, S. Y., Wong, W. A., O'Brien, H. E., Biron, V. L., Pires, J. C., Olmstead, R. G., Chase, M. W., y Rai, H. S. (2006). «Robust inference of monocot deep phylogeny using an expanded multigene plastid data set.» (pdf). Aliso (22): pp. 3-21. http://depts.washington.edu/phylo/OlmsteadPubs/Graham.2006.Aliso.pdf. Consultado el 25/02/2008.
- ↑ Hilu, K.; Borsch, T., Muller, K., Soltis, D. E., Soltis, P. S., Savolainen, V., Chase, M. W., Powell, M. P., Alice, L. A., Evans, R., Sauquet, H., Neinhuis, C., Slotta, T. A. B., Rohwer, J. G., Campbell, C. S., y Chatrou, L. W. (2003). «Angiosperm phylogeny based on matK sequence information.». American J. Bot. (90): pp. 1758-1766. http://www.amjbot.org/cgi/content/full/90/12/1758. Consultado el 25/02/2008.
- ↑ Pires, J. C.; Maureira, I. J., Givnish, T. J., Sytsma, K. J., Seberg, O., Petersen, G., Davis, J. I., Stevenson, D. W., Rudall, P. J., Fay, M. F., y Chase, M. W. (2006). «Phylogeny, genome size, and chromosome evolution of Asparagales.». Aliso (22): pp. 278-304. http://cat.inist.fr/?aModele=afficheN&cpsidt=18269727. Consultado el 25/02/2008.
- ↑ Rudall, P.; Furness, C. A., Chase, M. W., y Fay, M. F. (1997a). «Microsporogenesis and pollen sulcus type in Asparagales (Lilianae).». Canad. J. Bot. (75): pp. 408-430. http://cat.inist.fr/?aModele=afficheN&cpsidt=10769694. Consultado el 25/02/2008.
- ↑ a b Källersjö M; JS Farris, MW Chase, B Bremer, MF Fay, ,CJ Humphries, G Petersen, O Seberg, y K Bremer (1998). «Simultaneous parsimony jacknife analysis of 2538 rbcL DNA sequences reveals support for major clades of green plants, land plants, and flowering plants.». Pl. Syst. Evol. (213): pp. 259-287. http://www.springerlink.com/content/u864187n02112656/. Consultado el 25/02/2008.
- ↑ Fay, M. F. (2000). «Phylogenetic studies of Asparagales based on four plastid DNA regions.». En K. L. Wilson y D. A. Morrison. Monocots: Systematics and evolution. (Royal Botanic Gardens edición). Kollingwood, Australia: CSIRO. pp. 360-371.
- ↑ Soltis DE; PS Soltis, MW Chase, ME Mort, DC Albach, M Zanis, V Savolainen, WH Hahn, SB Hoot, MF Fay, M Axtell, SM Swensen, LM Prince, WJ Kress, KC Nixon, y JS Farris. (2000). «Angiosperm phylogeny inferred from 18S rDNA, rbcL, and atpB sequences.». Bot. J. Linn. Soc. (133): pp. 381-461. http://www.ingentaconnect.com/content/ap/bt/2000/00000133/00000004/art00380. Consultado el 25/02/2008.
- ↑ Stevenson, D. W.; Davis, J. I., Freudenstein, J. V., Hardy, C. R., Simmons, M. P., y Specht, C. D. (2000). «A phylogenetic analysis of the monocotyledons based on morphological and molecular character sets, with comments on the placement of Acorus and Hydatellaceae.». En Wilson, K. L. y Morrison, D. A.. Monocots: Systematics and evolution. (CSIRO Publ. edición). Collingwood, Australia. pp. 17-24.
- ↑ McPherson, M. A.; Graham, S. W. (2001). «Inference of Asparagales phylogeny using a large chloroplast data set. Abstract.». Botany 2001: pp. 126.
- ↑ Goldblatt, P. (1995). «The status of R. Dahlgren's orders Lliliales and Melanthiales.». En Rudall, P. J., Cribb, P. J., Cutler, D. F., y Humphries, C. J.. Monocotyledons: Systematics and evolution. (Royal Botanic Gardens edición). Kew. pp. 181-200.
- ↑ Stevenson, D. W.; Loconte, H. (1995). «Cladistic analysis of monocot families.». En Rudall, P. J., Cribb, P. J., Cutler, D. F.. Monocotyledons: Systematics and evolution. (Royal Botanic Gardens edición). Kew. pp. 543-578.
- ↑ Vinnersten, A.; Bremer, K. (2001). «Age and biogeography of major clades in Liliales». Amer. J. Bot. (88): pp. 1695-1703. http://www.amjbot.org/cgi/content/abstract/88/9/1695. Consultado el 25/02/2008.
- ↑ Soltis, D. E.; Soltis, P. F., Endress, P. K., y Chase, M. W. (2005). Phylogeny and evolution of angiosperms.. Sunderland, MA: Sinauer Associates.
- ↑ a b c Tamura, M. N. (1998). «Liliaceae». En K. Kubitzki. The families and genera of vascular plants, vol. 3, Monocotyledons: Lilianae (except Orchidaceae).. Berlin: Springer-Verlag. pp. 343-353.
- ↑ Hayashi, K.; Kawano, S. (2000). «Molecular systematics of Lilium and allied genera (Liliaceae): Phylogenetic relationships among Lilium and related genera based on the rbcL and matK gene sequence data.». Plant Species Biology 15: pp. 73-79. http://www.ingentaconnect.com/content/bsc/psb/2000/00000015/00000001/art00025;jsessionid=111qejj9ogz8u.alice?format=print. Consultado el 23/03/2008.
- ↑ a b Patterson, T. B.; Givnish, T. J. (2002). «Phylogeny, concerted convergence, and phylogenetic niche conservatism in the core Liliales: Insights from rbcL and ndhF sequence data.» (pdf). Evolution 56: pp. 233-252. http://www.botany.wisc.edu/givnish/Conc.converg.Liliales2002.pdf. Consultado el 23/03/2008.
- ↑ Conran, J. G. (1998). «Smilacaceae». En K. Kubitzki. The families and genera of bascular plants, vol. 6, Celastrales, Oxalidales, Rosales, Cornales, Ericales.. Berlin: Springer-Verlag. pp. 417-422.
- ↑ a b Fay, M. F.; Chase, M. W., Ronsted, N., Devey, D. S., Pillon, Y., Pires, J. C., Petersen, G., Seberg, O., y Davis, J. I. (2006). «Phylogenetics of Liliales: summarized evidence from combined analyses of five plstid and one mitochondrial loci.». Aliso (22): pp. 559-565.
- ↑ Rudall, P.; Stobart, K. L., Hong, . -P., Conran, J. G., Furness, C. A., Kite, G. C., y Chase, M. W. (2000). «Consider the lilies: systematics of Liliales.». En Wilson, K. L. y Morrison, D. A.. Monocots: Systematics and evolution. (CSIRO Publ. edición). Collingwood, Australia. pp. 347-359.
- ↑ Givnish, T. J.; Pires, J. C., Graham, S. W., McPherson, M. A., Prince, L. M., Patterson, T. B., Rai, H. S., Roalson, E. H., Evans, T. M., Hahn, W. J., Millam, K. C., Meerow, A. W., Molvray, M., Kores, P. J., O'Brien, H. E., Hall, J. C., Kress, W. J., y Sytsma, K. J. (2006). «Phylogeny of the monocots based on the highly informative plastid gene ndhF : Evidence for widespread concerted convergence». Aliso 22: pp. 28-51.
- ↑ Dahlgren, R. M.; Clifford, H. T., Yeo, P. F. (1985). The families of the monocotyledons. (Springer-Verlag edición). Berlín.
- ↑ Tamura, M. N. (1998). «Calochortaceae». En K. Kubitzki. The families and genera of vascular plants, vol. 3, Monocotyledons: Lilianae (except Orchidaceae).. Berlin: Springer-Verlag. pp. 164-172.
- ↑ Shinwari, Z. K.; Terauchi, R., Hteck, F. H., y Kawano, S. (1994). «Recognition of the New World Disporum section Prosartes (Liliaceae) based on the sequence data of the rbcL gene.». Taxon (43): pp. 353-366. http://links.jstor.org/sici?sici=0040-0262(199408)43%3A3%3C353%3AROTNWD%3E2.0.CO%3B2-U. Consultado el 25/02/2008.
- ↑ a b Kazuhiko Hayashi, Seiji Yoshida, Frederick H. Utech, Shoichi Kawano (2001). «Molecular systematics in the genus Clintonia and related taxa based on rbcL and matK gene sequence data». Plant Species Biology 16 (2) , 119–137 doi:10.1046/j.1442-1984.2001.00057.x. http://www.blackwell-synergy.com/doi/abs/10.1046/j.1442-1984.2001.00057.x. Consultado el 25/04/2008.
- ↑ Dahlgreen R. M. T., Clifford H. T. and Yeo P. F., 1985, The families of the monocotyledons: structure, evolution and taxonomy. Springer Verl., Stuttgart
- ↑ Ownbey, M. 1940. A monograph of the genus Calochortus. Ann. Missouri Bot. Gard. 27: 371–560.
- ↑ McDonald, H.P. A review of Calochortus. Herbertia 55:34-51; 2000
- ↑ Patterson, T. B., and Givnish, T. J., 2002, Phylogeny, concerted convergence, and phylogenetic niche conservatism in the core Liliales: Insights from rbcL and ndhF sequence data: Evolution, v. 56, p. 233–252.
- ↑ «Genera of Liliaceae» (en inglés). USDA, ARS, National Genetic Resources Program. Germplasm Resources Information Network - (GRIN) [Online Database].. National Germplasm Resources Laboratory, Beltsville, Maryland. (2001 en adelante). Consultado el 05-4-2008.
- ↑ a b flower bulbs at Aboutflowerbulbs.com
- ↑ Frankel, R. & Galun, E. (1977). Pollination mechanisms, reproduction, and plant breeding. Monographs on Theoretical and Applied Genetics no. 2. Springer-Verlag. Berlin. ISBN 0-387-07934-3.
Bibliografía
- Judd, W. S.; C. S. Campbell, E. A. Kellogg, P. F. Stevens, M. J. Donoghue (2007). «Liliaceae». Plant Systematics: A Phylogenetic Approach (3ª edición edición). Sunderland, Massachusetts: Sinauer Associates. ISBN 978-0-87893-407-2.
- Simpson, Michael G. (2005). «Liliaceae». Plant Systematics.. Elsevier Inc.. pp. 180. ISBN 0-12-644460-9 ISBN 978-0-12-644460-5.
Enlaces externos
 Wikimedia Commons alberga contenido multimedia sobre Liliaceae. Commons
Wikimedia Commons alberga contenido multimedia sobre Liliaceae. Commons Wikiespecies tiene un artículo sobre Liliaceae. Wikispecies
Wikiespecies tiene un artículo sobre Liliaceae. Wikispecies
- Watson, L.; Dallwitz, M. J.. «Liliaceae» (en inglés). The families of flowering plants: descriptions, illustrations, identification, and information retrieval. Versión: 25 de noviembre de 2008. Consultado el 15 de enero del 2009.
- Liliaceae de Chile en Chileflora











Wikimedia foundation. 2010.