- Glutamato deshidrogenasa
-
Glu deshidrogenasa (NAD (P)+) 1 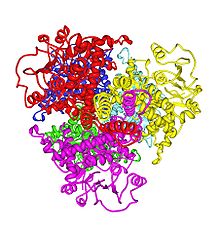
Estructura 3D de la GLUD humana. PDB 1L1F.HUGO 4335 Símbolo GLUD1 Símbolos alt. GLUD, GDH Datos genéticos Locus Cr. 10 q21.1-q24.3 Bases de datos Número EC 1.4.1.3 Entrez 2746 OMIM 138130 PDB 1L1F RefSeq NM_005271 UniProt P00367 Glu deshidrogenasa (NAD (P)+) 2 HUGO 4336 Símbolo GLUD2 Símbolos alt. GLUDP1 Datos genéticos Locus Cr. X q24-q25 Bases de datos Número EC 1.4.1.3 Entrez 2747 OMIM 300144 PDB 1L1F RefSeq NM_012084 UniProt P49448 La enzima Glutamato deshidrogenasa cataliza la reacción de oxidación del glutamato a 2-oxoglutarato desprendiéndose amoníaco.
- L-glutamato + H2O + NAD (P)+
 2-oxoglutarato + NH3 + NAD (P) H
2-oxoglutarato + NH3 + NAD (P) H
Hasta la fecha se han encontrado tres enzimas diferentes en la naturaleza:
- Glutamato deshidrogenasa (NAD (P)+), EC 1.4.1.3, llamada abreviadamente GLUD y utiliza tanto NADH como NADPH.
Todas estas glutamato deshidrogenasas conjuntanmente con la fenilalanina deshidrogenasa, leucina deshidrogenasa, y valina deshidrogenasa están estructuralmente y funcionalmente relacionadas. Un residuo conservado de lisina localizado en la región rica en glicina está implicado en el mecanismo catalítico.
Contenido
Glutamato deshidrogenasa (NAD (P)+) (GLUD)
La GLUD es una enzima, presente en la mayoría de los microbios y en la mitocondria de los eucariontes Se han caracterizado enzimas de bos taurus (bovinos), gallus gallus (pollo), drosophila melanogaster (mosca), zea mays (maíz), mus musculus (rata) y sus scrofa (cerdo).
El ser humano posee dos isozimas de la glutamato deshidrogenasa, la GLUD1 y la GLUD2. Ambas forman homohexámeros y su localización celular es la matriz mitocondrial.
GLUD1
La GLUD1 tiene un rol clave en el metabolismo del nitrógeno, del glutamato y de la homeostasis energética. Se expresa en altos niveles en el hígado, cerebro, páncreas y riñones, pero no en los músculos. Se piensa que en las células pancreáticas la GLUD1 participa en el mecanismo de secreción de insulina. En los tejidos nerviosos, en donde la concentración de glutamato es superior que en otros tejidos, la GLUD1 participa tanto en la síntesis como en el catabolismo del glutamato y quizás en la detoxificación del amoniaco. La GLUD1 puede estar involucrada en las reacciones del aprendizaje y de la memoria incrementando la producción del neurotransmisor glutamato.
Estructura
La GLUD1 es un homohexámero. Cada monómero tiene:
- Dominio N-terminal de unión del glutamato (Glu-BD), que está casi totalmente compuesto de filamentos β.
- Dominio de unión del NAD+ o NADP+ (NAD-BD).
- 48 residuos parecidos a una antena que se extienden desde la parte superior de cada NAD-BD. La antena consiste en una hélice ascendiente y un filamento descendiente que contiene una pequeña α-hélice que se dirige hacia el lado C-terminal del filamento.
EL NAD-BD se sienta sobre la parte superior del Glu-BD. El NAD-BD y el Glu-BD forman la ranura catalítica. Durante la unión del sustrato, el NAD-BD se mueve significantemente. Este movimiento tiene dos componentes, rotación alrededor del eje largo de una hélice en la espalda del NAD-BD, llamada hélice pivote y enrollamiento en la antena siguiendo las agujas del reloj. Una comparación de las conformaciones abierta y cerrada de la GLUD1 revela cambios en la hélice pequeña del filamento descendente de la antena, que parece que retrocede cuando la ranura catalítica se abre.[1] El cerramiento de una subunidad está asociado con la distorsión de la pequeña hélice del filamento descendente que es empujada hacia la antena de la subunidad adyacente.
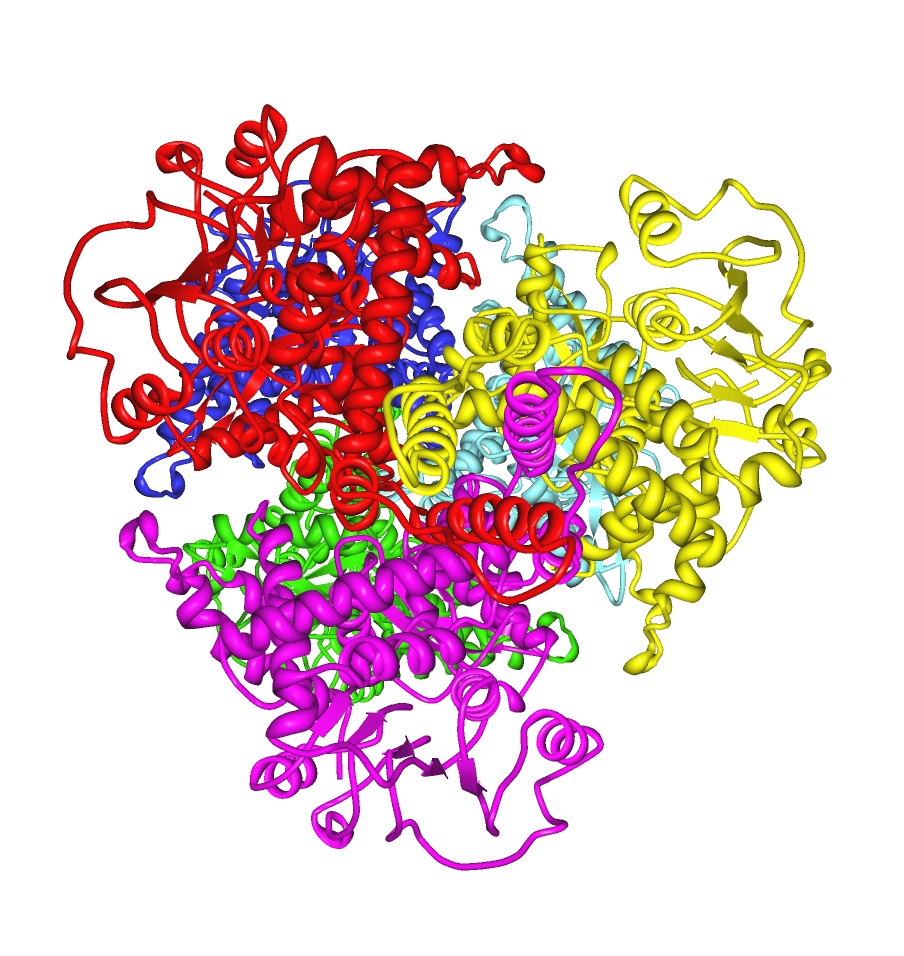
 Estructura de la GLD1
Estructura de la GLD1
Cada dominio esta coloreado diferente - Glu-BD, NAD (P)-BD, antena, la hélice pivote. Los reguladores alostéricos están mostrados como modelos de esferas. Esta estructura particular de la GLUD1 es una combinación de dos estructuras por rayos X - una con una GTP unida (1HWZ) y la segunda con un ADP unido (1NQT). Aunque no es real, esta estructura muestra la posición relativa de los efectores alostéricos cuando se unen a la GLUD1. El NADPH y el glutamato también se muestran.La estructura del núcleo del hexámero es una pila de dos trímeros. Los Glu-BD's de los monómeros son responsables principales de la construcción del núcleo. La posición relativa de los monómeros es tal que la rotación sobre la hélice pivote de cada monómero no está restringida. Las antenas de las tres subunidades de los trímeros sufren cambios conformacionales cada vez que la ranura catalítica se abre y se cierra. La antena sirve como medio de comunicación entre las subunidades durante la cooperación negativa y la regulación alostérica.
El estudio de las GLUD1 procedientes de diversas fuentes muestra que la antena probablemente evolucionó en los protistas antes de la formación de los sitios regulatorios para las purinas. Esto sugiere que hay alguna ventaja selectiva en la antena y que los animales evolucionaron nuevas funciones para la GLUD1 añadiendo la regulación alostérica.[2]
La GLUD1 puede formar largas fibras mediante la asociación de hexámeros. La polimerización no está relacionada con la actividad catalítica, pero probablemente tiene un papel importante como la formación de complejos multienzimáticos.
La GLUD1 tiene dos sitios de unión para co-enzimas: uno en el NAD-BD que es capaz de unirse con el NAD+ o NADP+ y participa directamente en el proceso catalítico; y el segundo, que tiene una función reguladora y está situado debajo justo de la hélice pivote, puede unir ADP, NAD+ o NADH, pero no puede unir correctamente el NADPH.
Unión de ADP y GDP
El ADP se une a la glutamato deshidrogenasa detrás del NAD-BD, justo debajo de la hélice pivote; el segundo sitio de unión para coenzimas. La parte adenosina se une en un agujero hidrofóbico con los grupos ribosa fosfato apuntando hacia la hélice pivote. El ADP también se puede unir al segundo sitio (inhibitorio) del NADH causando activación.
La unión del GTP es antagonizada por el fosfato y el ADP pero es sinérgica con la unión del NADH en el sitio alostérico no catalítico. La mayoría de los contactos entre el GTP y la enzima se efectúan a través de la parte trifosfato. Al sitio de unión del GTP se le considera el sensor que apaga la enzima cuando la célula está en un situación rica en energía. El GTP se une en la unión entre el NAD-BD y la antena.[3] [4] Aunque la mayoría de las interacciones GLUD1-GTP se efectúan a través de interacciones con los fosfatos β y γ, hay interacciones específicas con el Glu346 y la Lys343 que favorecen a la guanosina sobre la adenosina. En la conformación abierta, el sitio de unión del GTP está distorsionado de tal forma que no puede unirse con el GTP.[1]
Mecanismo de reacción
La GLUD1 cataliza la deaminación oxidativa del glutamato a 2-oxoglutarato e ion amonio (NH4) usando NAD+ o NADP+ como cofactor. La reacción se desarrolla con la transferencia de un ion hidruro desde el carbono α del glutamato al NAD (P)+, formándose 2-iminoglutarato, que es hidrolizado a 2-oxoglutarato y NH4. El equilibrio de reacción en condiciones standard favorece la formación de glutamato frente al NH4 (ΔG≈30 kJ/mol). Por esta razón, se piensa que la enzima tiene un papel importante en la detoxificación del amoniaco; esta posición de equilibrio ayuda a mantener la concentración de NH4 baja.
Regulación
Cuando la GLUD1 está altamente saturada por los ligandos (sustratos) de los sitios activos, se forma un complejo inhibitorio en el sitio activo: NAD (P) H-Glu en la reacción de deaminación oxidativa a pH alto, y NAD (P)+-2-oxoglutarato en la reacción de aminación reductiva a pH bajo. La GLUD1 asume su configuración basal en la ausencia de efectores alostéricos. Los reguladores alostéricos de la GLUD1 (ADP, GTP, Leu, NAD+ y NADH) ejercen sus efectos cambiando la energía requerida para la transformación de la ranura catalítica en abierta o cerrada, en otras palabras desestabilizando o estabilizando respectivamente los complejos inhibitorios. Los activadores no son necesarios para la función catalítica de la GLUD1, ya que es activa en la ausencia de estos compuestos (estado basal). Se ha sugerido que la GLUD1 asume en su estado basal una configuración (ranura catalítica abierta) que permite la actividad catalítica con independencia de si sus sitios alostéricos son funcionales. La regulación de la GLUD1 es de particular importancia biológica, ejemplificado por las observaciones de que las mutaciones regulatorias de la GLUD1 están asociadas con manifestaciones clínicas en los niños.
El ADP es uno de los dos principales activadores de la enzima (siendo el NAD+ el segundo). Actúa desestabilizando los complejos inhibitorios y anulando la cooperación negativa. En la ausencia de sustratos y con el ADP unido, la ranura catalítica está en la conformación abierta y los hexámeros de GLUD1 forman largos polímeros con más interacciones (PDB 1NQT) que en la forma inhibida. Esto es consistente con el hecho que el ADP promueve la agregación en solución. Cuando la ranura catalítica se abre, el residuo Arg-516 rota hacia abajo sobre los fosfatos del ADP.[3] La abertura de la ranura catalítica está correlacionada con la distancia entre la Arg-516 y los fosfatos del ADP. De esta forma, el ADP activa la GLUD1 facilitando la abertura de la ranura catalítica que disminuye la afinidad por los productos y facilita la liberación de los mismos.[5] [1] se ha sugerido que la inhibición por una alta concentración de ADP es debida a la competencia en el sitio activo entre el ADP y la parte adenosina de la coenzima.
La concentración de ATP tiene complejos efectos sobre la actividad de la GLUD1. Una baja concentración de ATP significa inhibición de la enzima. La afinidad del ATP por el sitio del GTP es 1000 veces inferior a la del GTP, ya que las interacciones de los fosfatos β y γ son determinantes para la unión en el sitio del GTP. Si la concentración de ATP es media, se produce una activación de la enzima, mediada a través del sitio efector del ADP. Por último, se produce una inhibición de la proteína si la concentración de ATP es alta. Se ha sugerido que esto es debido al mismo efecto que tiene una concentración alta de ADP, es decir a la competencia en el sitio activo entre el ATP y la parte adenosina de la coenzima.[6]
El GTP inhibe la enzima en un amplio abanico de condiciones incrementando la afinidad de la GLUD1 por el producto de reacción. El GTP hace que la etapa limitadora de la velocidad de reacción sea la liberación del producto. El GTP actúa manteniendo la ranura catalítica en una conformación cerrada estabilizando por tanto los complejos inhibitorios. Los efectos del GTP en la GLUD1 no son solamente locales a la subunidad a la que está unida, la antena juega un papel importante en comunicar esta inhibición a las otras subunidades.
El NAD (P)(H) se puede unir a la enzima en un segundo sitio en cada subunidad. El NAD (H) se une 10 veces mejor que el NADP (H), y la forma reducida mejor que la oxidada. Se ha sugerido que la unión de la coenzima reducida inhibe la enzima y que la unión de la coenzima oxidada la activa, aunque los efectos aún no están claros.
Otros reguladores de la enzima son:
- La leucina (Leu) activa la GLUD1 uniéndose a ésta en cualquier sitio, quizás directamente en la ranura catalítica.
- El fosfato y otros aniones estabilizan la GLUD1. Estudios estructurales recientes muestran que las moléculas de fosfato se unen al sitio del GTP.[3]
A modo de resumen:
Regulador Activación GLUD1 Inhibición GLUD1 ADP En bajas y medias concentraciones. En altas concentraciones. ATP En medias concentraciones. En bajas y altas concentraciones. GTP Siempre. NAD (P) H / NAD (P)+ Coenzima oxidada. Coenzima reducida. Leucina Siempre. Fosfato Siempre. Gen de la GLUD1
 Estrucura exón/intrón del gen GLUD1.El esquema de colores es el siguiente: Glu-BD, NAD (P)-BD, antena, la hélice pivote
Estrucura exón/intrón del gen GLUD1.El esquema de colores es el siguiente: Glu-BD, NAD (P)-BD, antena, la hélice pivoteEl gen humano de la GLUD1 contiene 13 exones y está localizado en el cromosoma 10. Hay evidencias de que el gen GLUD1 ha sido retro-puesto en el cromosoma X en donde codifica la GLUD2.
Enfermedades
La deficiencia en GLUD1 es causa del Síndrome de hiperinsulinismo-hiperamonemia (HHS), caracterizado por una elevada velocidad de oxidación de glutamato a alfa-cetoglutarato que estimula la secreción de insulina en las células beta pancreáticas, mientras que no existen una detoxificación adecuada del amonio en el hígado.
GLUD2
La GLUD2 es la segunda glutamato deshidrogenasa humana que se expresa principalmente en la retina, testículos y en un menor nivel en el cerebro. Es importante para reciclar el principal neurotransmisor excitatorio, el glutamato, durante la neurotransmisión. El gen de la GLUD2 se encuentra en el cromosoma X y al contrario que la GLUD1 es un gen sin intrones.[7]
Glutamato deshidrogenasa
La glutamato deshidrogenasa es una proteína NAD-específica que cataliza la deaminación oxidativa del glutamato. Se ha caracterizado en seres vivos como la Achlya klebsiana, Bacteroides fragilis, Bacillus subtilis, Clostridium difficile, Clostridium symbiosum, Neurospora crassa, Peptostreptococcus asaccharolyticus, Porphyromonas gingivalis, Schizosaccharomyces pombe y Staphylococcus aureus. La longitud de las enzimas de estos seres varía entre 400 y 1100 aminoácidos encontrándose dos familias diferentes: la de ~450 AA y la de ~1050 AA. La glutamato hidrogenasa se asocia en homotetrámeros u homohexámeros.
Glutamato deshidrogenasa (NADP+)
La glutamato deshidrogenasa (NADP+) es una proteína NADP-específica que cataliza la deaminación oxidativa del glutamato. Se ha caracterizado en seres vivos como la Halobacterium salinarium, Agaricus bisporus, Botryotinia fuckeliana, Chlorella sorokiniana, Corynebacterium efficiens, Corynebacterium glutamicum, Debaryomyces occidentalis, Escherichia coli, Emericella nidulans y Giardia lamblia. La longitud de las enzimas de estos seres vivos varía entre los 400 y 550 aminoácidos, asociándose en homohexámeros. La enzima de la C. sorokiniana puede ser un homo o heterohexámero de subunidades alfa y beta expresadas por el mismo gen.
Referencias
- ↑ a b c The structure of apo GLUD details subunit communication and allostery. JMB. 2002; 318: 765-77; Abstract
- ↑ Structural studies on ADP activation of mammalian GLUD and the evolution of regulation. Biochemistry. 2003; 42: 3446-56; Abstract;
- ↑ a b c Structures of btGLUD complexes elucidate the mechanism of purine regulation. JMB. 2001; 307: 707-20; Abstract;
- ↑ The structure of btGLUD provides insights into the mechanism of allostery. Structure Fold Des. 1999; 7: 769-82; Abstract;
- ↑ Effects of ADP on btGLUD: diethyl pyrocarbonate modification. Biochemistry. 1980; 19: 6057-61; Abstract;
- ↑ Expression, purification and characterization of GLUD1 allosteric regulatory mutations. Biochem J. 2002; 363(Pt 1): 81-7; Texto libre
- ↑ Novel GLUD expressed in neural and testicular tissues and encoded by an X-linked intronless gene. JBC. 1994; 269: 16971-76; Texto libre (PDF - 3.2 MB);
Enlaces externos
- Glutamato deshidrogenasa (NAD (P)+) en NiceZyme (en inglés).
- Glutamato deshidrogenasa en NiceZyme (en inglés).
- Glutamato deshidrogenasa (NADP+) en NiceZyme (en inglés).
- GLUD1 en Wikipedia inglesa.
Categorías:- Genes del cromosoma 10
- Genes del cromosoma X
- Oxidorreductasas
- L-glutamato + H2O + NAD (P)+
Wikimedia foundation. 2010.