- Liquen
-
 Liquen sobre una pared de cemento.
Liquen sobre una pared de cemento.
 Liquen.
Liquen.Los líquenes son organismos que surgen de la simbiosis entre un hongo llamado micobionte y un alga o cianobacteria llamada ficobionte.
De acuerdo con el carácter de esa asociación, se pueden distinguir numerosos tipos estructurales de líquenes: desde el más simple, donde hongo y alga se unen de forma casual, al más complejo, donde el micobionte y el ficobionte han dado lugar a un talo morfológico muy diferente a aquel al que pertenecen por separado, y donde el alga se encuentra formando una capa bajo la protección del hongo.[1]
Los líquenes son organismos excepcionalmente resistentes a las condiciones ambientales adversas y capaces, por tanto, de colonizar muy diversos ecosistemas. La protección frente a la desecación y la radiación solar que aporta el hongo y la capacidad de fotosíntesis del alga confieren al simbionte características únicas dentro de los seres vivos. La síntesis de compuestos únicamente presentes en estos organismos, las llamadas sustancias liquénicas permiten un mejor aprovechamiento de agua, luz y la eliminación de sustancias perjudiciales.
Contenido
Taxonomía
El grupo de organismos que denominamos líquenes es un grupo polifilético, es decir proveniente de multitud de ancestros diferentes que ha evolucionado hacia un mismo patrón partiendo de relaciones diferentes; aun así no existe una clasificación para este grupo completamente aceptada por todos los expertos. La clasificación de Ozenda y Clauzade (publicada en 1970) atiende en primer lugar al tipo de hongo que forma la simbiosis, de este modo se diferencian tres clases: Ascolichenes, Basidiolichenes e Hypholichenes; según sea el hongo un ascomiceto, un basidiomiceto o un deuteromiceto, respectivamente. Dentro del primer grupo se diferencian, a su vez, dos subclases Pyrenolichenes y Discolichenes según posean peritecios o apotecios. Los ascolichenes constituyen el 96% de los líquenes siendo muy pocos los basidiolíquenes.[2]
Hoy día la posición taxonómica de los líquenes se encuentra en proceso de investigación y es muy extendida la tendencia a agruparlos dentro de los hongos ascomycotas en la clase Lecanoromycetes que es la que concentra la mayoría de los micosimbiontes conocidos; esta clasificación sin embargo deja fuera de la definición de liquen a aquellos formados por basidiomicotas y oomicotas. Otra tendencia incluye a todos los líquenes conocidos dentro de la División Mycophycota del reino Fungi, pero considerando que la única característica que permite la permanencia de esta división es la formación de simbiosis, no sus características citológicas, genéticas o filogénicas. Las clasificaciones más actuales basadas en estudios genéticos ofrecen más en profundidad los vínculos existentes entre las diferentes familias de hongos aunque quedan varias de ellas cuya posición filogenética no está del todo clara y continuamente aparecen enmiendas y rectificaciones para estos modelos.[3] [4] Es posible que los estudios genéticos que se realizan en la actualidad acaben con esta disyuntiva confirmando la polifilesis del grupo o identificando el origen común de estos organismos.[5]
Historia evolutiva
Los restos fósiles de líquenes son extraordinariamente escasos, es conocido en el mundo de la paleobotánica que el registro fósil, incompleto, no puede mostrar en absoluto la realidad de la flora de la época a la que pertenece, es por ello que se hace necesario deducir a partir de los escasos rastros conservados y a partir de la filogenia cuándo aparecen muchos de los grupos vegetales y en nuestro caso la simbiosis entre un alga y un hongo.
Aún con esto, el yacimiento de Rhynie Chert ha dado un ejemplo de liquen fósil de extraordinario valor científico pues sitúa a este grupo en era devónica pudiéndose considerar por tanto a este fósil como el más antiguo de los conocidos para el grupo.[6]
Consideraciones coevolutivas y estudios filogenéticos que relacionan distribuciones actuales y movimientos continentales han sugerido que el estilo de vida liquénico es muy antiguo. Parece probable que muchas de las familias, géneros, y en algunos casos especies actuales evolucionaron en tiempos del Pérmico/Triásico, hace unos 190-280 millones de años, a partir de unas cuantas especies previas. Existen teorías que mantienen que pudieran ser los líquenes las primeras especies en colonizar el medio terrestre, teoría demasiado controvertida y con pruebas aún muy poco consistentes.[7]
En el intento de presentar en conjunto los órdenes de la subdivisión Ascomycotina en relación con su biología, se encontró que el orden Peltigerales ocupa una posición más o menos central. También se observó, que este orden presenta un significado particular en la evolución de los ascomycetes, ya que incluye algunos géneros que son esencialmente terrícolas, aunque capaces de propagarse sobre cortezas rugosas y árboles; esto es, se presentan generalmente en hábitats primarios, los cuales han existido antes del aumento de las fanerógamas. Las cianobacterias pasan por ser uno de los primeros organismos fotosintéticos; presentes en la tierra desde el Precámbrico, es por ello que este tipo de fotobionte pudo estar presente para los hongos capaces de asociarse a él desde el principio evolutivo del grupo.[8] [9]
La simbiosis
La base de la simbiosis es la toma de nutrientes por parte del hongo desde el alga; para ello en casi todos los líquenes estudiados se ha encontrado alguna forma de penetración del hongo dentro de las células algales, la que se logra por medio de haustorios. Se diferencian dos tipos de haustorios u órganos de penetración del hongo: intracelulares e intramembranosos.[10] En los líquenes cruatáceos o crustosos (que forman costras) y en algunas formas más altamente estructuradas las penetraciones son generalmente intracelulares donde los haustorios penetran en el protoplasto de la capa gonidial (donde se encuentra el alga); la pared de estos haustorios es más delgada que la del resto de las hifas de forma que les resulta más sencillo penetrar en la célula vegetal. En los líquenes morfológicamente más evolucionados los haustorios son intramembranosos; en estos casos penetran la pared de la capa gonidial pero no el citoplasma, quedando una invaginación en la pared del alga. El liquen obtiene su alimento a partir de las sustancias sintetizadas por el alga a través de la fotosíntesis; en este proceso se sintetiza el hidrato de carbono denominado ribitol que es transferido al hongo mediante difusión, en el interior de la hifa este ribitol es modificado a manitol que no es transferible desde el hongo al alga.[11] De este modo el micobionte se asegura su alimento y el alga sigue sintetizando. Cuando el fotobionte es una cianobacteria, el hidrato de carbono sintetizado es la glucosa que el hongo modifica también a manitol.[12] [13] El alga por su parte consigue del hongo la protección necesaria frente a la desecación, un incremento de su capacidad de absorción de agua gracias a las características de las hifas del hongo. En definitiva la simbiosis permite al alga o cianobacteria colonizar ecosistemas donde debido a un clima extremo no podría desarrollarse por sí sola.
Los metabolitos de los líquenes suelen ser una mezcla de los producidos por el alga para su propio funcionamiento y los producidos por el hongo. Son pocos los casos en los que se producen sustancias propias para el liquen que ninguno de los biontes podría producir por sí mismo; entre estas sustancias se encuentran las sustancias liquénicas, un conjunto muy heterogéneo de productos específicos de los líquenes producidos por el hongo, muchas de ellas denominadas ácidos liquénicos. Se conocen alrededor de 200 tipos de sustancias liquénicas aunque las investigaciones añaden constantemente nuevos tipos, por lo que se piensa que muchas son exclusivas de una sola especie, queriendo esto decir que es la simbiosis misma la que las produce. Vale aclarar que estas sustancias no aparecen en el córtex inferior ni en la capa gonidial y en casi todos los casos se presentan en forma de minúsculos cristales y granulaciones dispuestas en la superficie de las hifas. La función de las sustancias liquénicas no está del todo clara; se piensa que pueden actuar como método disuasorio para herbívoros o como protección ante bacterias y otros patógenos. Es posible también que jueguen un papel importante en la absorción de agua o aumentando la permeabilidad de la membrana de las algas permitiendo de ese modo la entrada de metabolitos al interior celular. Si se sabe que actúan como protección frente a diversos contaminantes, a las radiaciones UV e incluso a la radioactividad. Algunas de ellas son simplemente desechos de la actividad celular cotidiana del organismo simbionte.[14] Estos metabolitos liquénicos son la base de muchos estudios sobre taxonomía en estos organismos.[15]
Tipos de biontes
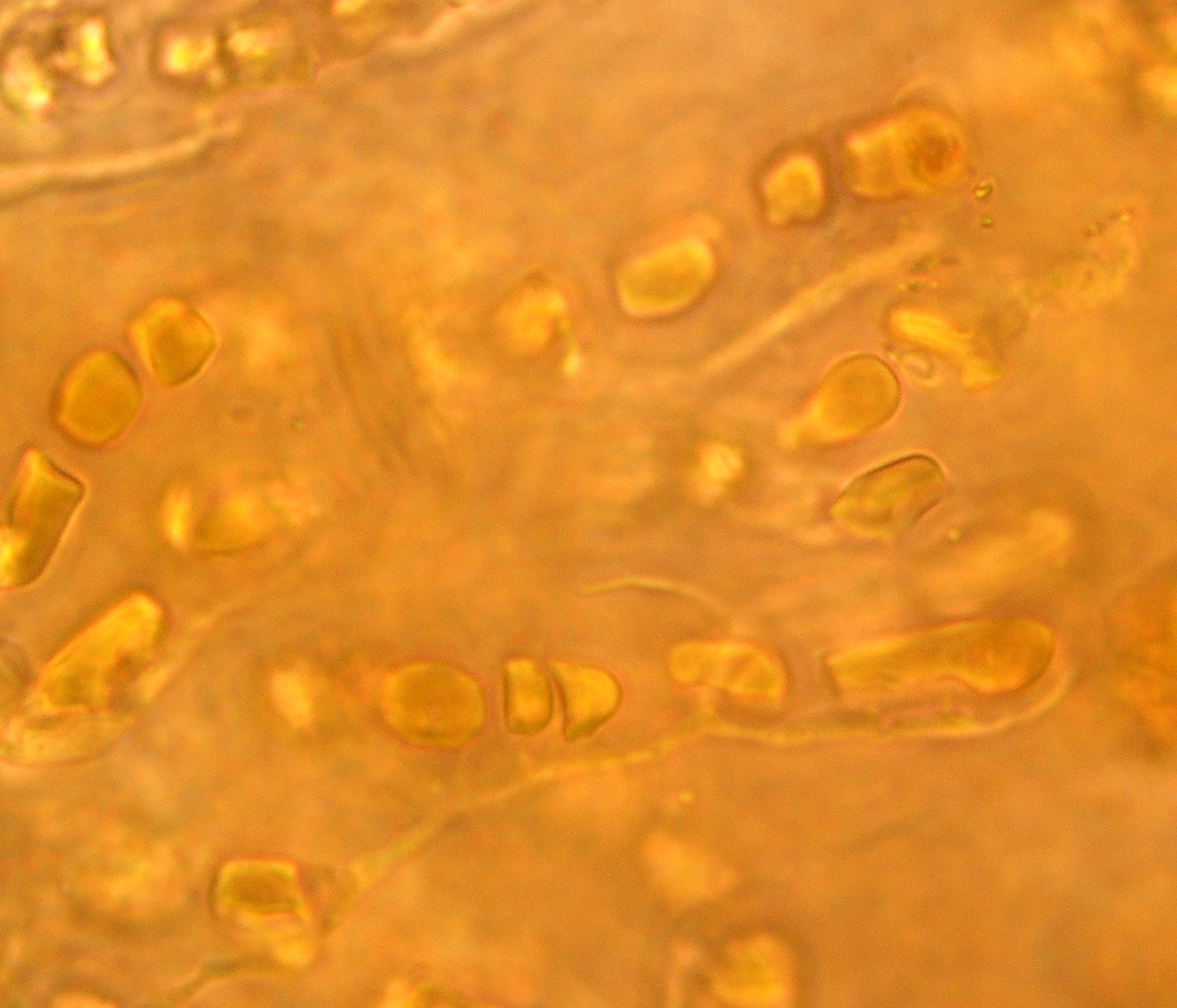
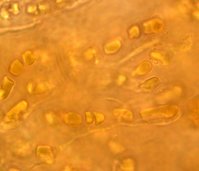 La cianobacteria Hyella caespitosa junto a hifas de un hongo en el liquen Pyrenocollema halodytes.
La cianobacteria Hyella caespitosa junto a hifas de un hongo en el liquen Pyrenocollema halodytes.Puede decirse que la formación de simbiosis, sobre todo mutualistas por parte de los hongos con todo tipo de fotobiontes es una característica que ha dado una enorme ventaja evolutiva a las especies que las forman, al menos esto puede desprenderse de los datos que sobre especies que forman algún tipo de simbiosis poseemos, así del total de 64.200 especies de hongos, alrededor de un treinta por ciento (19.000 especies) han optado por este tipo de asociación, más de un ocho por ciento forman simbiosis como micorrizas y un veintiuno por ciento forman las asociaciones liquénicas.
Entre los mayores grupos de hongos, la cantidad de especies implicadas en asociaciones liquénicas es enorme, por ejemplo el 98% de esos hongos pertenece a la subdivisión Ascomycota, pero de los cuarenta y seis órdenes conocidos de hongos dieciséis forman líquenes y de ellos sólo seis forman únicamente líquenes, es decir que no se conocen como hongos de vida libre. Existen relativamente pocos hymenomycetes liquenizados, sirvan de ejemplo Dictyonema, Multiclavula, y Omphalina, un liquen mastigomicete (Geosiphon pyriformes) y alrededor de cincuenta géneros de hongos conidiales.[16]
A priori puede afirmarse que desde el punto de vista evolutivo la formación de simbiosis liquénicas es una estrategia evolutiva muy favorable y que se encuentra en constante evolución en algunos hongos, por ejemplo en Hymenomycetes o Helotiales, aunque parece que también está perdiéndose en otras, como en Caliciales y Lecanorales.
Cuando el micobionte es un ascomiceto, se denomina a la simbiosis con el alga ascolíquen, que presenta las fructificaciones típicas de los ascomicetos, en caso de que se trate de peritecios se denominan pirenolíquenes (pirenomicetos) y si se trata de apotecios, discolíquenes (discomicetos); en el caso de que el micobionte sea un basidiomiceto se denomina al simbionte basidioliquen.
El ficobionte suele ser una clorofita, más raramente una cianofita; a veces se presentan asociaciones triples, como en el género Lobaria, un ascolíquen, con una clorofita como ficobionta, que, sin embargo, presenta unas estructuras especiales, llamadas cefalodios, en las que se produce una asociación con una cianoficea (Nostoc).[17] Estos cefalodios se desarrollan en aquellos simbiontes liquénicos en los que se hace necesaria una simbiosis extra con una cianofícea a ser el alga simbionte principal incapaz de fijar el nitrógeno atmosférico necesario para el hongo.[18] Debido a su naturaleza dual, los líquenes presentan características tanto del hongo como del alga, pese a presentar características propias y particulares.
Fotobiontes
Actualmente se conocen aproximadamente cuarenta géneros de algas y cianobacterias que actúan como fotobiontes en simbiosis liquénica. De ellos tres géneros son los más frecuentes, Trebouxia, Trentepohlia y Nostoc, las dos primeras algas verdes y la tercera cianobacterias.[19]
Los fotobiontes eucariotas son conocidos como ficobiontes mientras que los fotobiontes cianobacterias son conocidas como cianobiontes; la inmensa mayoría de los ficobiontes son algas verdes (phyllum Chlorophyta) que poseen clorofila a y b y sólo dos géneros (del phyllum Heterokontophyta) poseen clorofila a y c. La transferencia metabólica entre fotobiontes y micobiontes depende en gran medida del tipo de fotobionte presente de este modo cuando se trata de un alga verde el carbohidrato compartido posee grupos alcohol (ribitol) mientras que en líquenes con cianobacterias es glucosa.
La identificación de los fotobiontes cianobacterias en talos liquénicos es a veces imposible debido a que la morfología del talo de la cianobacteria cambia debido a la presencia del micobionte, por ello las formas filamentosas como ocurre con el género Dichothrix suelen estar tremendamente deformadas; sólo los filamentos ramificados de géneros como Stigonema pueden identificarse en el talo liquénico.[20]
Las cianobacterias tampoco muestran todas las fases de su ciclo vital cuando forman parte de un liquen, ello dificulta aún más su identificación como sucede en géneros como Chloroccidiopsis y Myxosarcina; la observación de todas las formas vitales de este tipo de organismo suele ser primordial para su correcta taxonomización a nivel de especie, se hace necesario en el trabajo con ficobiontes el aislamiento del alga y su posterior cultivo en vida libre. El tipo de reproducción vegetativa es también un carácter tremendamente importante en la taxonomía, así especies con heterocistes (como Nostoc) aumentan la frecuencia de producción de éstos cuando se encuentran en simbiosis en comparación a cuando viven de forma libre; también el tamaño de las células suele ser mayor es especies en simbiosis como ocurre con Gloeocapsa en los líquenes del género Lichinella o Peccaria, este incremento suele ser interpretado como resultado de la presencia de gran cantidad de haustorios del hongo en el interior de la célula algal y a la estrecha unión de éstos con el vegetal.
En algas verdes la organización del talo es siempre muy simple cuando actúan como fotobiontes, sólo son conocidas formas estructurales en coco, sarcinas y filamentos donde las estructuras filamentosas suelen ser muy reducidas en tamaño.
La identificación del alga verde que forma el liquen es con frecuencia más fácil que cuando se trabaja con cianobacteria, al menos en multitud de ocasiones no es necesario aislar el alga y cultivarla para poder identificarla al menos hasta nivel de género, para afinar hasta especie sí son esenciales los cultivos debido a que la morfología de los cloroplastos y varias etapas del ciclo de vida algal no modificados o están ausentes en la simbiosis con el hongo.
Micobiontes
Los hongos formadores de líquenes son en gran parte de los casos simbiontes obligados y no son capaces de vivir aislados en el medio; sólo prosperan cuando encuentran un fotobionte adecuado, en cultivo aislado muestran su forma imperfecta pudiendo producir esporas asexuales pero prácticamente nunca producen estructuras reproductoras organizadas como los hongos no liquenizados ni estructura del talo que se asemeje al del liquen.
Actualmente no existen pruebas que permitan afirmar que los hongos que forman parte de la simbiosis liquénica sean diferentes morfológicamente de los de vida libre.[21] Es cierto que se ha encontrado en algunos grupos de Ascomycotas unos orgánulos esféricos, de naturaleza proteínica y función desconocida que no se encuentran en hongos libres y que algunos autores han pretendido otorgar funciones relacionadas con la simbiosis pero las últimas investigaciones apuntan que más bien puede tratarse de orgánulos propios de la especie de Ascomycota formadora de líquenes en la que se encontró y no debido a la simbiosis con el alga.[22]
Estos cuerpos concéntricos han sido con posterioridad localizados en el citoplasma de micelios, células ascógenas, parafisis y todo tipo de células en hongos patógenos de plantas y saprobios sobre todo de ambientes secos por lo que es probable que tengan una función de almacenamiento de sustancias o resistencia a la desecación, necesaria eso sí, en los hongos liquenizados.
La principal diferencia entre hongos liquenizados y no liquenizados es por supuesto el tipo de nutrición que llevan a cabo. Muchos hongos aparecen en la naturaleza sólo formando parte de líquenes aunque en cultivo se han podido aislar del alga y han sido capaces de sobrevivir.
Los cultivos de estos hongos formadores de líquenes de forma aislada han dado casi en todos los casos un fenotipo muy diferente al expresado cuando forman la simbiosis, una gran proporción de estos hongos han presentado un talo característico formado por masas celulares aglutinadas con crecimiento filamentoso sólo en la periferia; sin embargo alguno de los grupos más interesantes, como por ejemplo los Perigerales, sólo están capacitados para crecer cuando forman parte de un liquen, de este modo cultivos de esporas de diversas especies de este grupo han sido cultivadas aisladas del alga y aunque han germinado su micelio ha sido incapaz de desarrollarse, únicamente tras la adición de un fotobionte compatibles el ahora liquen ha crecido y con él el hongo.[23]
Organización del talo liquénico
Se denomina talo homómero a aquel en el que fotobionte y micobionte se encuentran distribuidos de forma uniforme.[24] Son los líquenes gelatinosos los que principalmente poseen estructura homómera de su talo. Estos líquenes son capaces de absorber más agua que los líquenes no gelatinosos en relación a su peso seco. Esto hace que el intercambio de gases sea muy limitado en estos organismos ya que cabe la posibilidad de que el talo se encuentre saturado de agua y por lo tanto la difusión del dióxido de carbono (CO2) se vea dificultada enormemente. Por ello que en estos líquenes la presencia del CO2 es un factor limitante para la fotosíntesis. Recientemente se han descrito diversos métodos de concentración de CO2 que permiten su acúmulo en el talo para posteriores usos por el fotobionte.[25]
Los talos heterómeros son aquellos en los que fotobionte y micobionte ocupan diferentes estratos dentro del liquen. Se divide el talo en varias capas, por una parte aparece un córtex superficial, de hifas muy apretadas de hongo donde por lo general nunca se encuentran rastros del alga. A continuación aparece la llamada capa gonidial, con hifas laxas mezcladas con células algales, es la región donde se produce la fotosíntesis por parte del alga y la interacción de ésta con el hongo se hace más patente por la presencia de los haustorios. Por último la médula con hifas poco apretadas del hongo.
Por último se habla de estructura filamentosa a la constituida por filamentos de una clorofita o una cianofita revestidos por hifas.
Morfología del talo en el simbionte liquénico.
Son escasos los ejemplos de líquenes cuya morfología viene determinada por el fotobionte (ocurre por ejemplo en los géneros Coenogonium, Ephebe, Cystocoleus o Racodium), en la mayoría de los casos es el micobionte el que marca las pautas de crecimiento para el simbionte.[9] El desarrollo de cierto tipo de talo es importante para conocer las relaciones que se establecerán en el simbionte, por ello tradicionalmente se ha dividido al grupo en la morfología de su talo en líquenes crustáceos, foliáceos y fruticulosos, a pesar de esta clasificación existen otros biotipos posibles en líquenes, como los llamados gelatinosos que algunos autores, sin embargo, incluyen en los tipos anteriores.
Talo crustáceo
 Liquen crustáceo.
Liquen crustáceo.Los llamados talos crustáceos son aquellos que crecen fuertemente unidos al sustrato, hasta el punto de que es imposible separarlos de él sin destruirlo. Las características del talo de este tipo de líquenes les permite sobrevivir en ambientes muy extremos y en superficies expuestas de roca. Poseen organización tanto homómera como heterómera sobre todo en los márgenes de aquellos que posean areolas grandes o en especies intermedias con los líquenes foliosos. No poseen corteza inferior; su crecimiento es marginal pudiendo muchas veces solaparse diversos individuos y formar características estructuras a parches; la forma más simple de organización está presente en géneros como Lepraria en los que las hifas del micobionte envuelven pequeños grupos de alga sin posibilidad de diferenciarlos, el talo posee apariencia pulverulenta como afectado por lepra, razón por la cual el género toma el nombre; en el género de líquenes epífitos Vezdaea el fotobionte se encuentra organizado en pequeños gránulos o soredios de apenas un milímetro de diámetro.
 Liquen crustáceo escamoso.
Liquen crustáceo escamoso.La estructura de los líquenes endolíticos (que viven en el interior de microfisuras de rocas) o endofloeódicos (bajo la cutícula de hojas de plantas) es mucho más compleja; en muchos casos existe diferenciación entre el córtex superior y el resto del talo; diversos géneros como Buellia y Lecidea incluso se desarrollan estratificación del talo en ausencia del sustrato; este último género sitúa al fotobionte en la médula y puede extenderse hasta dos milímetros en el interior de los granos arenosos de las rocas en la que viven llegando a introducirse muchos más como sucede en la especie Lecidea sarcogynoides de Sudáfrica donde diversos individuos han penetrado hasta 9.6 milímetros de profundidad cada cien años.
Los líquenes crustáceos, que viven unidos fuertemente a la superficie de las rocas, pueden presentar muy diversa morfología. De este modo encontramos especies con los márgenes no limitados, esbozados que apenas se diferencian del sustrato. Líquenes con bordes bien delimitados, de color más claro o más oscuro que el resto del individuo y bien diferenciados del medio. Talos figurados, lobulados radialmente y con los bordes laxamente unidos al sustrato pudiendo incluso separarse de él. Por último los talos areolados poseen una división en su cara superior con numerosos surcos que delimitan porciones o areolas, los surcos dejan ver la zona más interna del individuo, de color oscuro.[26]
Los líquenes epilíticos son los más abundantes dentro de los líquenes crustáceos, su talo suele encontrarse perfectamente limitado en su margen o con límites imprecisos. Un talo areolado es aquel en el que el simbionte se distribuye en múltiples porciones (areolas) de formas poligonales, en la estación seca cada una de la areolas de este organismo son perfectamente distinguibles mientras que en la estación húmeda no lo son; las areolas se desarrollan a partir de un talo primario por extensión hacia toda su circunferencia. El talo más complejo dentro de los crustáceos es el llamado escamoso donde las areolas crecen hasta llegar a separarse parcialmente del sustrato formando las características escamas que dan nombre al fenotipo.
Talo folioso
 Liquen folioso.
Liquen folioso.Los líquenes foliosos son aquellos en los que el talo se encuentra parcialmente despegado del sustrato y no en tan íntima relación con él como en los anteriores. Los talos pueden ser homómeros o heterómeros. Lo más usual es que posean organización dorsiventral, distinguiéndose entre zonas ventrales y dorsales. Dentro de este tipo de líquenes existe una enorme diversidad en cuanto a formas, organización y tamaños.
Los líquenes laciniados son aquellos que poseen la estructura típica de los líquenes foliosos; se adhieren al sustrato en casi toda su extensión y poseen en gran parte de las especies lóbulos cuya distribución en el talo es de los más variada, radial, alterna, etc. En algunas especies los lóbulos pueden encontrarse inflados por un mayor crecimiento de la médula. Son los líquenes que alcanzan mayores tamaños dentro del grupo y presentan un amplio abanico de colores, de consistencia y de formas, son ejemplo de ellos los géneros Xanthoparmelia, Physcia y Solorina. Se llama a un liquen umbilicado cuando posee un talo circular con un único anclaje al sustrato en el centro como ocurre en Umbilicaria.
Un tipo muy particular de líquenes foliosos crecen en desiertos y poseen un interesante movimiento higroscópico, en épocas de sequía son capaces de enrollarse sobre sí mismos para mostrar la menor cantidad posible de superficie y evitar de ese modo la desecación exponiendo su superficie inferior formada por hifas del hongo, además en estado enrollado son capaces de ser transportados por el viento, normalmente hasta lugares sombríos como bases de piedras o matorrales a la espera de la llegada de humedad, esto ocurre en especies como Xanthomaculina convoluta o Chondropsis semivirdis.[27]
Talo fruticuloso
 Liquen fruticuloso.
Liquen fruticuloso.El talo de los líquenes fruticulosos es alargado, cilíndrico o muy estrecho en todos los casos asemejándose a una cabellera, poseen por lo general un único punto de unión al sustrato quedando el resto del organismo lejos de él; pueden ramificarse, a veces muy profusamente, poseen crecimiento apical o intercalar y pueden ser macizos o huecos en el caso de los homómeros y aplanados los heterómeros. Sin embargo existen excepciones a esta morfología general, así Sphaerophorus melanocarpus posee simetría dorsiventral aunque la anchura de los lóbulos es muy reducida. El tamaño de estos líquenes es muy variable según la especie, por ejemplo el género Usnea crece varios metros mientras otras especies apenas lo hacen unos milímetros. La unión al sustrato se lleva cabo mediante estructuras de fijación especiales que en algunas especies degenera en la madurez dejando al individuo libre del sustrato. Ejemplos de este tipo de líquenes son los pertenecientes a los géneros Cladonia, Stereocaulon o Roccella.[28]
Talos heteromórficos
Algunos géneros desarrollan dos tipos de morfología en su talo así Baeomyces o Cladonia poseen una parte del talo fruticuloso y otra parte, por supuesto la unida al sustrato escamoso. Son los llamados talos dimórficos; en ellos se diferencia entre un talo horizontal o talo primario adherido al sustrato y otro vertical o secundario portador de los cuerpos fructíferos. Es posible que en estado adulto se pierda el talo primario como ocurre en Cladina.[29] En algunas especies el talo secundario está formado por tejido carpogénico, quiere decir esto que forma parte del cuerpo fructífero mientras que en otras especies el desarrollo del cuerpo fructífero se lleva a cabo en el extremo del talo secundario no siendo él mismo parte de ellos.
Reproducción
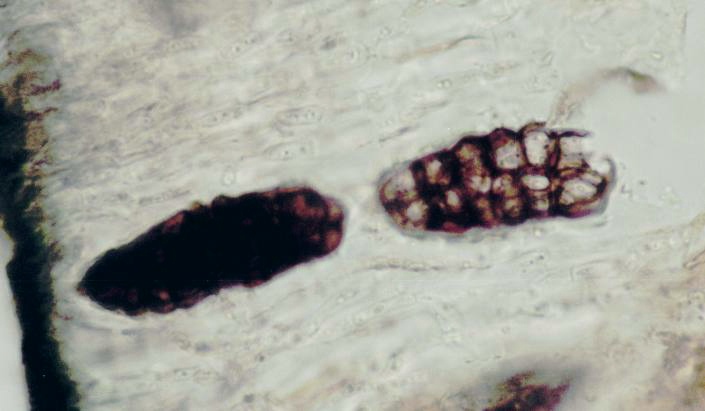
 Apotecios en la superficie de un liquen.
Apotecios en la superficie de un liquen. Fotografía de dos esporas de Rhizocarpon grande (Flörke ex Flotow) Arnold tomadas mediante un microscopio óptico con un aumento de 1000x. El ejemplar fotografiado es un espécimen de herbario colectado en Condado Hancock , Maine, Estados Unidos (11 de julio de 2005).
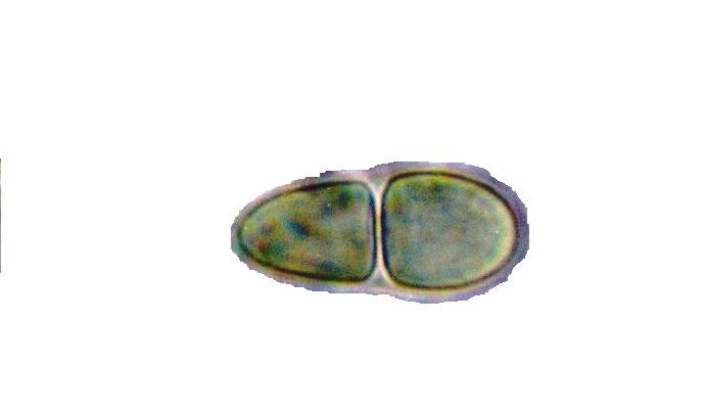
Fotografía de dos esporas de Rhizocarpon grande (Flörke ex Flotow) Arnold tomadas mediante un microscopio óptico con un aumento de 1000x. El ejemplar fotografiado es un espécimen de herbario colectado en Condado Hancock , Maine, Estados Unidos (11 de julio de 2005). Fotografía de una espora de Rhizocarpon hochstetteri, mostrando un septo. Ejemplar colectado en el Condado Washington, Maine, Estados Unidos el 9 de junio de 2001. Aumento: 1000x.
Fotografía de una espora de Rhizocarpon hochstetteri, mostrando un septo. Ejemplar colectado en el Condado Washington, Maine, Estados Unidos el 9 de junio de 2001. Aumento: 1000x.Los líquenes pueden reproducirse asexualmente a partir de porciones de talo con representantes de los dos biontes en la denominada fragmentación talina o a partir de estructuras especializadas llamadas soredios(que pueden a su vez estar agrupados en los denominados soralios) e isidios.
Un soredio es una agrupación de hifas del hongo rodeando a unos pocos elementos algares que se desprende de la superficie del talo liquénico para ser diseminado bien por el viento, bien por salpicaduras de lluvia, esta estructura carece por completo de organización interna.[30] Los isidios por el contrario se encuentran estructurados de la misma manera que el talo liquénico; son porciones de talo que se desarrollan en la superficie conservando la estructura en capas y córtex y que pueden desprenderse con facilidad. Estos tipos de reproducción asexual son los únicos procesos mediante los cuales se disemina la estructura completa del liquen y no sólo uno de sus componentes. Ha de advertirse que estas estructuras son creadas exclusivamente por el hongo al ser éste el único componente que necesita de la simbiosis para su supervivencia, o al menos el que tiene más ventaja en la unión. La ventaja evolutiva que implica la formación de estas estructuras pone de manifiesto la bondad de la simbiosis para el micobionte y su necesidad de asegurarse un organismo fotosintético para su supervivencia.[31]
El resto de las estructuras reproductoras formadas en el simbionte no involucran a los dos biontes. Debido probablemente a limitaciones impuestas por el micobionte que aún no se conocen el elemento fotobiótico es incapaz de reproducirse por sí mismo mientras forma parte de la simbiosis. El hongo por su parte es capaz de reproducirse asexual y sexualmente según las características propias del grupo al que pertenezca, ascomicete, basidiomicete u otro. Los componentes reproductores fúngicos son capaces de diseminarse en busca de un alga al que asociarse, que será la misma especie con la que previamente formaba la simbiosis u otra diferente dependiendo de la especialización del hongo.
Las esporas sexuales formadas en ascomicotas se encuentran característicamente en peritecios y apotecios; en líquenes estas estructuras pueden estar formadas exclusivamente por el hongo o tener parte de la capa algar participando de ellas, en ambos casos las estructuras producidas en el himenio se diseminan en busca en un nuevo fotobionte o desarrollan un hongo de vida libre, salvo en hongos que son incapaces de vivir fuera de la simbiosis.
El hongo también puede desarrollar estructuras reproductoras asexuales por su cuenta; son los llamados picnidios. Este tipo particular de esporas que se producen en gran cantidad es capaz de permanecer en el medio durante mucho tiempo a la espera de encontrar el alga o ciantofita adecuada con la que asociarse, como ocurre con las esporas sexuales.[32]
Ecología
Estos organismos son colonizadores primarios en casi todos los ecosistemas conocidos, su capacidad de adaptación a medios con escasos nutrientes hace que sean capaces de desarrollarse tempranamente y comenzar la formación de suelo para la posterior llegada de otros organismos vegetales.[33] Los líquenes son organismos muy específicos respecto al sustrato y a las condiciones del medio en el que se desarrollan. Es posible encontrar simbiontes liquénicos en medios extremadamente hostiles para la vida como zonas polares o desérticas donde las características que les aporta la simbiosis permiten su desarrollo. Según un experimento realizado por la Agencia Europea del Espacio durante 2005 dos especies de líquenes antárticos fueron capaces de sobrevivir en el espacio sin ningún tipo de protección.[34] Esta gran capacidad de supervivencia ha permitido que las diversas especies de simbiontes liquénicos hayan colonizado y prosperado en prácticamente todos los ecosistemas terrestres.
Existen varias especies de líquenes marinos correspondientes a varios fila de hongos Ascomicotas y que poseen en una gran parte de los casos al alga Trebouxia como fotobionte. Estos líquenes tienen como hábitat principal la zona intermareal donde la batida de las olas impide el crecimiento de muchas algas, de este modo la simbiosis beneficiaría al alga ya no tanto en cuanto al almacenaje de agua sino respecto a la protección mecánica. Es posible reconocer en estas comunidades una zonación vertical del medio donde se desarrollan los organismos, de esta manera existen especies que se encuentran siempre sumergidas y otras que sólo lo están en los momentos más altos de pleamar.[35]
Los líquenes son utilizados como indicador biológico de la calidad del aire debido principalmente a su susceptibilidad a la presencia de dióxido de azufre en el medio. Los talos jóvenes son mucho más sensibles a esta contaminación ambiental que aquellos plenamente desarrollados. La producción de soredios se ve notablemente reducida en medios contaminados por lo que la propagación de estos organismos se reduce notablemente. El dióxido de azufre es el principal responsable de la acidificación del agua de lluvia (Lluvia ácida) necesaria para el crecimiento de estos organismos, la respuesta del talo frente a este tipo de contaminación es la creación de sustancias liquénicas hidrófobas y la reducción de la superficie expuesta a la lluvia de modo que la fotosíntesis se ve reducida y con ella el crecimiento aparte de la cantidad de agua utilizable.[36] La recuperación, sin embargo, de estos organismos ha resultado ser espectacular una vez que las condiciones ambientales vuelven a ser normales. Así, según estudios realizados tras la eliminación de gran parte de los contaminantes ambientales de la ciudad de Londres a finales del siglo XX los líquenes han vuelto progresivamente a colonizar aquellos hábitats que les eran apropiados.[37]
Referencias
- ↑ Nash, Thomas H. (1996). Lichen biology.. Cambridge University Press. ISBN 0-521-45974-5.
- ↑ P. Ozenda & G. Clauzade (1970) Les Lichens. Étude biologique et flore illustré. Ed. Masson et Cie., París.
- ↑ Modelo de Lumbsch y Huhndorf en 2007.
- ↑ Modelo ofrecido en 2007 en Mycological reseach.
- ↑ A Gargas, PT DePriest, M Grube, and A Tehler, Multiple origins of lichen symbioses in fungi suggested by SSU rDNA phylogeny. Science, Vol 268.
- ↑ Líquenes fósiles de Rhynie chert.
- ↑ http://russian.china.org.cn/english/scitech/128654.htm Líquenes fósiles que parecen demostrar la colonización del medio terrestre encontrados en China].
- ↑ Daniel S. Heckman et al. First land plants and fungi changed earth's climate, paving the way for explosive evolution of land animals, new gene study suggests, Science, agosto de 2001 [1]
- ↑ a b David L. Hawksworth, Interacciones hongo-alga en simbiosis liquénicas y liquenoides Anales del Real Jardín Botánico de Madrid 46
- ↑ Lichen biology Thomas H. Nash. Página 2
- ↑ J. W. Deacon, Modern Mycology p.111
- ↑ Vernon Ahmadjian, The Lichen Symbiosis p.100
- ↑ A química dos liquens en Quimica Nova volumen 22 n.1 São Paulo 1999.
- ↑ Ecología química en hongos y líquenes. Francisco J Toledo Maranten et al.
- ↑ Aplicación de técnicas analíticas e interpretación de las variaciones químicas en líquenes. Esteban Manrique Reol.
- ↑ R. S. Mehrotra y K. R. Aneja, An Introduction to Mycology p. 706
- ↑ Martha Chaparro de Valencia, Jaime Aguirre Ceballos, Hongos liquenizados p.80
- ↑ María Isabel Martínez Moreno (1999) Taxonomía del género Peltigera Willd. (Ascomycetes liquenizados) en la península Ibérica y estudio de sus hongos liquenícolas, p.34 en Monografías del Real Jardín Botánico de Madrid, CSIC
- ↑ Ahmadjian, Vernon (1993). The Lichen Symbiosis. Pag. 30. John Wiley & Sons, Inc.. ISBN 0-471-57885-1.
- ↑ Ahmadjian, Vernon (1993). The Lichen Symbiosis. Pag. 56.. John Wiley & Sons, Inc.. ISBN 0-471-57885-1.
- ↑ Thomas H. Nash, Lichen Biology p.24
- ↑ H. B. Griffiths1 y A. D. Greenwood, The concentric bodies of lichenized fungi en Archives of Microbiology Volumen 87, Nº4 pp.285 a 302 diciembre de 1972 ISSn 0302-8933
- ↑ B. Büdel y C. Scheidegger en Lichen Biolohy de Thomas H. Nash, p.65
- ↑ Martha Chaparro de Valencia, Jaime Aguirre Ceballos, Hongos liquenizados p.35
- ↑ B. Büdel y C. Scheidegger en Lichen Biolohy de Thomas H. Nash, p.88
- ↑ B. Büdel y C. Scheidegger en Lichen Biology p.39
- ↑ Thomas H. Nash, Lichen Biology p.43
- ↑ Martha Chaparro de Valencia, Jaime Aguirre Ceballos, Hongos liquenizados p.32
- ↑ Martha Chaparro de Valencia, Jaime Aguirre Ceballos, Hongos liquenizados p.34
- ↑ Eichorn, Susan E., Evert, Ray F., and Raven, Peter H. 2005. Biology of Plants. New York (NY):W.H. Freeman and Company. 289 p.
- ↑ Vernon Ahmadjian, The Lichen Symbiosis p.25
- ↑ Vernon Ahmadjian, The Lichen Symbiosis p.64
- ↑ R. S. Mehrotra, K. R. Aneja, An Introduction to Mycology p.708
- ↑ Noticia Universia
- ↑ Fungaldiversity
- ↑ Ecored200
- ↑ Lichen recolonization in London's cleaner air. C. I. Rose and D. L. Hawksworth in Nature 289, 289 - 292
Bibliografía
- Galun, Margalith (1988). Handbook of Lichenology. CRC. ISBN 0-8493-3582-5.
- Chaparro de Valencia, Martha (2002). Hongos liquenizados.. UNIBIBLOS. ISBN 958-701-241-0.
- Seckbach, Joseph (2002). Simbiosis: Mechanisms and model Systems.. Dordrecht: Kluwer Academic Publishers. ISBN 1-4020-0189-4.
- Nash, Thomas H. (1996). Lichen biology.. Cambridge University Press. ISBN 0-521-45974-5.
- Ahmadjian, Vernon (1993). The Lichen Symbiosis.. John Wiley & Sons, Inc.. ISBN 0-471-57885-1.
- Volkmar et al. (2004). Guía de campo de líquenes, musgos y hepáticas.. Ediciones Omega. ISBN 84-282-1266-X.
- Surindar Paracer y Vernon Ahmadjian (2000). Symbiosis: An Introduction to Biological Associations. Oxford University Press. ISBN 0-19-511807-3.
- R. S. Mehrotra, K. R. Aneja (1990). An Introduction to Mycology. New Age Publishers. ISBN 81-224-0089-2.
- Deacon, J. W. (1997). Modern Mycology. Blackwell Publishing. ISBN 0-632-03077-1.
- Hawksworth, David L. (1988). «Interacciones Hongo-Alga en simbiosis liquénicas.». Anales del Jardín Botánico de Madrid VOLUMEN (46). 235-247. http://www.rjb.csic.es/pdfs/Anales_46(1)_235_247.pdf?PHPSESSID=.
Enlaces externos
Categorías:- Líquenes
- Especies indicadoras
- Simbiosis






Wikimedia foundation. 2010.