- Poaceae
-
 Poáceas
Poáceas
TrigoClasificación científica Reino: Plantae División: Angiospermae Clase: Monocotyledoneae Subclase: Commelinidae Orden: Poales Familia: Poaceae Barnhart (= Gramineae Juss., nom. cons.) Subfamilias y tribus - Anomochlooideae Potztal
- Anomochloeae
- Streptochaeteae
- Aristidoideae Caro
- Arundinoideae Burmeist.
- Bambusoideae Luerss.
- Centothecoideae Soderstr.
- Centotheceae
- Thysanolaeneae
- Chloridoideae Beilschm.
- Danthonioideae Barker & Linder
- Danthonieae
- Ehrhartoideae Link
- Micrairoideae Pilger
- Eriachneae
- Micraireae
- Panicoideae Link
- Pharoideae L.G. Clark & Judziewicz
- Phareae
- Pooideae Bentham
- Puelioideae L.G. Clark, Kobay., Mathews, Spangler & Kellogg
- Guaduelleae
- Puelieae
Las gramíneas o poáceas (Poaceae Barnhart) son una familia de plantas herbáceas, o muy raramente leñosas, perteneciente al orden Poales de las monocotiledóneas (Liliopsida). Con más de 670 géneros y cerca de 10.000 especies descritas, las gramíneas son la cuarta familia con mayor riqueza de especies luego de las compuestas (Asteraceae), las orquídeas (Orchidaceae) y las leguminosas (Fabaceae); pero, definitivamente, es la primera en importancia económica global.[1] De hecho, la mayor parte de la dieta de los seres humanos proviene de las gramíneas, tanto en forma directa (granos de cereales y sus derivados, como harinas y aceites) o indirecta (carne, leche y huevos que provienen del ganado y las aves de corral que se alimentan de pastos o granos). Es una familia cosmopolita, que ha conquistado la mayoría de los nichos ecológicos del planeta, desde las zonas desérticas hasta los ecosistemas de agua salada, y desde las zonas deprimidas y anegadizas hasta los sistemas montañosos más altos. Esta incomparable capacidad de adaptación está sustentada en una enorme diversidad morfológica, fisiológica y reproductiva y en varias asociaciones mutualísticas con otros organismos, que convierten a las gramíneas en una fascinante familia, no solo por su importancia económica, sino también por su relevancia biológica.
Descripción
Hábito de crecimiento
En general son hierbas, si bien pueden ser leñosas —como los bambúes tropicales—, cespitosas, rizomatosas o estoloníferas. Por la duración de su ciclo de vida pueden ser anuales, bienal o perennes. Las gramíneas anuales, como es lógico suponer, se reproducen una sola vez durante su ciclo vital —el caso del trigo o de la avena, por ejemplo—. Las especies perennes, en cambio, pueden reproducirse varias veces —en general anualmente— o una sola vez. En el primer caso se denominan iteróparas —la mayoría de las especies de pastos, por ejemplo— y, en el segundo caso, semélparas —como es el caso de las diferentes especies bambúes—.
Tallo
Tienen tallos cilíndricos a elípticos en su sección transversal, articulados, llamados ordinariamente cañas, en general con nudos macizos y entrenudos huecos (pero pueden ser totalmente macizos como en el caso del maíz y algunos bambúes). Los nudos son algo más gruesos que los entrenudos y en ellos nacen las hojas y las yemas. Los entrenudos son a veces algo achatados en la zona donde se desarrollan las ramificaciones. Un poco más arriba del nudo existe un meristema intercalar en forma de anillo que determina el alargamiento del tallo. En algunos géneros existen de 2 a 6 nudos muy próximos entre sí (los cuales se denominan nudos compuestos), cada uno de los cuales lleva su correspondiente hoja. En Cynodon dactylon, por ejemplo, los nudos están en grupos de a dos por lo que las hojas parecen opuestas. En general los entrenudos basales son más cortos que los superiores; cuando hay varios nudos basales muy próximos, las hojas parecen arrosetadas, es decir, se disponen de forma tal que simulan formar una roseta basal de hojas. Los principales tipos de tallos en las gramíneas son los siguientes:
- Tallos aéreos:
- Tallos erectos o ascendentes, generalmente con entrenudos cortos en la base y gradualmente más largos hacia el ápice. Pueden ser simples o ramificados.
- Tallos rastreros: que crecen recostados sobre el suelo, arraigando en los nudos; frecuentemente presentan nudos compuestos, como en Stenotaphrum y Cynodon. Cuando los tallos rastreros originan nuevas plantas en cada nudo se denominan estolones.
- Tallos flotantes, los cuales flotan en el agua gracias a los entrenudos huecos o a la presencia de aerénquima.
- Tallos subterráneos:
- Rizomas: existen dos tipos de rizomas en las gramíneas. Un primer tipo de rizomas cortos y encorvados, de crecimiento definido, que generan nuevas plantas junto a la mata original contribuyendo a aumentar su diámetro, como por ejemplo en Arundo donax y Spartina densiflora. El otro tipo de rizoma es largo, de crecimiento indefinido que propaga la planta a cierta distancia de la mata original. Ejemplos de este segundo tipo son Sorghum halepense y Panicum racemosum.
- Pseudobulbos, son engrosamientos de los entrenudos basales envueltos por sus vainas foliares. Son poco frecuentes en las gramíneas, algunos ejemplos son Amphibromus scabrivalvis y Phalaris tuberosa.[2] [3]
Hojas
Poseen hojas de disposición alterna, dísticas, compuestas típicamente de vaina, lígula y limbo. La vaina rodea apretadamente al tallo, sus márgenes se superponen pero no se fusionan entre sí (sólo ocasionalmente pueden ser encontradas formando un tubo). La lígula es un pequeño apéndice membranoso, o raramente un grupo de pelos (tricomas), situado en la zona de unión del limbo con la vaina, en la parte adaxial. El limbo (o lámina) es simple, usualmente lineal, con nerviación paralela. Puede ser aplanado o a veces enrollado en un tubo, puede ser continuo con la vaina o poseer pecíolo. Además de esta descripción, es necesario abordar la variabilidad que se puede hallar para cada uno de éstos órganos:
 Lígula membranosa en avena (Avena sativa).
Lígula membranosa en avena (Avena sativa).
 Aurículas en trigo (Triticum aestivum).
Aurículas en trigo (Triticum aestivum).- Profilo: es la primera hoja de cada innovación, generalmente está reducida a una vaina membranosa, con 2 carenas conspicuas, sin lígula ni lámina, que protege a la innovación.
- Vaina: la vaina nace en un nudo y envuelve a la caña, pudiendo ser más corta o más larga que el entrenudo. Frecuentemente la vaina esta hendida hasta su base, si bien sus bordes se superponen enrollándose alrededor de la caña, pero en muchas especies está parcial o totalmente soldada por sus bordes (ejemplo, en Bromus) e incluso la lígula puede formar un anillo continuo alrededor de la caña, como ocurre en Melica y en algunas especies de Poa. Las nervaduras de la vaina son numerosas y uniformes, si bien en especies con cañas comprimidas las vainas pueden presentar una carena conspicua.
- Lígula: generalmente membranosa, pero en algunas tribus, por ejemplo en las Eragrostídeas, Arundinóideas y Panicóideas, está formada por una banda de pelos o cilias, o bien no existe. En algunos géneros de Tritíceas y Festúceas a los costados de la lígula hay dos aurículas que abrazan a la caña.
- Pseudopecíolo: en muchas Bambúseas existe una contracción entre la lámina y la vaina que semeja un pecíolo; frecuentemente este pseudopecíolo está articulado con la lámina y es persistente. También hay pseudopecíolos en el género Pharus y en Setaria palmifolia.
- Lámina: usualmente la lámina es lineal o lanceolada, entera en el margen y con nervaduras paralelas. En los géneros tropicales aparecen láminas oblongas hasta anchamente elípticas, como en Pharus y Olyra. La lámina de Neurolepis (Bambusoideae) puede tener hasta 4 metros de largo. Cuando las láminas son anchas y planas, como en Zea y en Sorghum, hay una nervadura central conspicua. En cambio en las hojas lineales, estrechas, las nervaduras son más o menos iguales entre sí. En estos casos suele haber láminas que se pliegan longitudinalmente (se dicen conduplicadas) o que se enroscan (convolutas). Esta característica se aprecia perfectamente en la primera prefoliación. Permite, además, identificar a ciertas especies. Por ejemplo, las hojas conduplicadas son típicas de Stenotaphrum secundatum, Bromus brevis, Dactylis glomerata, Axonopus compressus, Poa lanuginosa, entre otras. Las hojas convolutas se pueden apreciar en Lolium multiflorum, Bromus unioloides, Paspalum dilatatum, entre varios otros ejemplos. En algunos casos la lámina se modifica haciéndose gruesa y subulada, como en Sporobolus rigens.
- Epidermis: los caracteres epidérmicos de la hoja y de las brácteas de las gramíneas tienen gran importancia en sistemática y con frecuencia sirven para diferenciar determinadas subfamilias o tribus. En general, la disposición de las células epidérmicas es diferente en las caras superior e inferior de la lámina. También es distinta la disposición sobre los haces vasculares (se llaman zonas costales) y entre dichos haces (zonas intercostales). Las células de la epidermis de las gramíneas se pueden agrupar en 5 categorías:
- Células silíceas, no poseen coloración, se reconocen por su refringencia o brillo particular ya que refractan la luz de un modo diferente al resto de las células epidérmicas debido a que poseen el lumen (el interior de la célula) ocupado por sílice. Pueden ser redondas, alargadas longitudinalmente, en forma de hacha de doble filo, en forma de tibia o contraídas en su parte central.
- Células suberosas, son células cortas, muertas a la madurez, con la pared celular suberificada.
- Células exodérmicas que comprenden los elementos que sobresalen de la superficie de la epidermis, los que incluyen: pelos unicelulares, bicelulares o pluricelulares; pelos capitados, pelos rígidos y punzantes (aguijones) más o menos silicificados y papilas (células con prominencias cónicas) muy cortas.
- Estomas, formados por 2 tipos de células: las células oclusivas o de cierre, en forma de tibia, alargadas y engrosadas en sus extremos, las cuales rodean al orificio por donde se realiza el intercambio gaseoso o poro estomático, y las células anexas, semilunares, rectangulares o trapezoidales.
- Células largas con paredes lisas u onduladas y células buliformes incoloras que forman bandas en el fondo de surcos sobre la cara superior de la lámina. Ciertos elementos epidérmicos son comunes a todas las Gramíneas, como las células largas, las suberosas, los pelos unicelulares y los aguijones. Otros son característicos y particulares de ciertos grupos.
- Anatomía foliar: la anatomía de la hoja de las gramíneas tiene gran importancia en sistemática. Existen dos tipos de anatomía extremas:
- Tipo Festucoide, con la vaina exterior de los haces vasculares (llamada vaina parenquimática ) poco diferenciada, con cloroplastos, y la vaina interior (denominada vaina mestomática) bien desarrollada y fuertemente engrosada, sin clorofila; además el clorénquima está formado por células sin invaginaciones o lóbulos, no está ordenado en ninguna forma particular. El parénquima incoloro no está presente. Este tipo de anatomía es la típica de las Festúceas y ciertas tribus tropicales como las Oríceas.
- Tipo Panicoide. La vaina mestomática está apenas desarrollada o falta por completo, mientras que la vaina parenquimática está muy desarrollada. En este tipo de anatomía el clorénquima tiende a ordenarse en forma radial alrededor de los haces vasculares, con células lobuladas. El parénquima incoloro está presente. Este tipo anatómico, característico de las Paníceas, Eragrósteas, Clorídeas y otras tribus tropicales, se denomina anatomía de Kranz y es propio de las gramíneas C4. En contraste, el tipo anatómico festucoide es propio de las gramíneas C3. Además, existen tipos intermedios como por ejemplo en las Bambúseas, donde tanto la vaina parenquimática como la mestomática están muy desarrolladas. Las diferencias en la anatomía de las hojas están asociadas con vías fotosintéticas diferentes. La Neurachne minor, Steinchisma decipiens (= Panicum decipiens), S. hians (= S. milioides) y en S. spathellosum (= S. schenckii).[2] [3]
Macollos
Los macollos o macollas son la unidad estructural de la mayoría de las especies de gramíneas. Se forman a partir de las yemas axilares o secundarias del meristema basal del eje principal. Cada uno de estos brotes secundarios o macollos inician su aparición cuando las plantas presentan entre dos y tres hojas. Cada uno de ellos, luego de producir sus primeras hojas, genera su propio sistema radicular. La suma o adición de macollos es lo que conforma la estructura y la forma de una planta de gramínea. Cuando las gramíneas se hallan en estado vegetativo producen continuamente nuevos macollos y hojas. Cada macollo, a su vez, comenzará en su momento a producir nuevos macollos.
Espiguilla
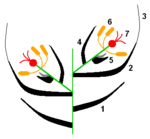 Esquema de una espiguilla: 1. Glumas; 2: Lemma; 3: Arista; 4: Pálea; 5: Lodículas; 6: Androceo; 7: Gineceo.
Esquema de una espiguilla: 1. Glumas; 2: Lemma; 3: Arista; 4: Pálea; 5: Lodículas; 6: Androceo; 7: Gineceo. Detalle de una espiguilla de gramínea, se observan las glumas, la lemma y la pálea.
Detalle de una espiguilla de gramínea, se observan las glumas, la lemma y la pálea.La inflorescencia elemental de las gramíneas es una pequeña espiga formada por una o más flores sentadas o sésiles sobre un raquis articulado, a menudo brevísimo, llamado raquilla y protegido por brácteas estériles denominadas glumas. Este tipo de inflorescencia recibe el nombre de espiguilla. Las flores pueden ser hermafroditas o unisexuales y presentan un perianto rudimentario de 2 ó 3 piezas, las lodículas o glumélulas. Estas lodículas son los órganos que, al ponerse turgentes, determinan la apertura del antecio o casilla floral durante la floración, permitiendo que se expongan los estigmas plumosos y los estambres. Los antecios están formados por la lemma, adherida a la raquilla, y la pálea insertada sobre el eje floral que nace sobre la raquilla en la axila de la lemma y soporta a los órganos florales propiamente dichos. La lemma (o glumela inferior) tiene forma de quilla, puede ser mútica o aristada, y abraza a la pálea con sus bordes. La arista nace en la extremidad de la lemma o en su dorso. La pálea o glumela superior es lanceolada, binervada y es como una tapa que encierra a la flor. Las glumas están insertadas sobre la raquilla, una más abajo que la otra. Todos estos elementos son muy variables por lo que es conveniente analizarlos por separado.
- Pedicelo. Es el pedúnculo que sostiene a la espiguilla que puede ser más o menos largo o estar completamente ausente, en ese caso las espiguillas son sésiles.
- Raquilla, o eje de la inflorescencia. Suele ser ondulada llevando en cada ángulo un antecio. La raquilla puede estar articulada con el pedicelo por debajo de las glumas (que entonces caen con la espiguilla) o por encima de las glumas (que, entonces, son persistentes). La raquilla puede ser tenaz o fragmentarse en artejos a la madurez de los frutos.[4] A veces la raquilla es persistente, desprendiéndose los frutos con las glumelas (como en el caso de Eragrostis bahiensis). También puede prolongarse en forma de arista glabra o velluda más allá del antecio superior, como en Deyeuxia.
- Glumas. Típicamente son dos, alternas, herbáceas, membranosas o papiráceas, ovadas o lanceoladas, contraídas lateralmente y más o menos carenadas,[5] o redondeadas, casi planas por compresión dorsiventral de la espiguilla (Paspalum). Pueden llevar una o varias nervaduras y ser múticas, mucronadas o aristadas en el ápice. En las Oríceas las glumas son rudimentarias o están ausentes. En cambio, en muchas Paníceas hay aparentemente 3 glumas, siendo la superior en realidad una lemma estéril; a veces hay una sola gluma como en los géneros Monerma y Lolium.
- Antecios. Los antecios (casilla floral, en griego) pueden ser desde uno hasta varios. Están formados por dos brácteas, las glumelas, que encierran a la flor. En algunos géneros los antecios se desprenden con un fragmento de la raquilla, frecuentemente cubierto de pelos que forma el antopodio o callus (Aristida, Stipa). En otros géneros con glumas caducas con el antecio, hay un callus formado por el ápice del pedicelo (ejemplo, en Heteropogon). La glumela inferior está generalmente más desarrollada y se denomina lemma; es la bráctea en cuya axila se desarrolla la flor; la lemma es ovada o lanceolada; comprimida lateral o dorsalmente, con una o varias nervaduras, aguda u obtusa, mútica o con una o más aristas apicales o dorsales. La glumela superior o pálea es el profilo situado entre la flor y la raquilla; generalmente es menor que la lemma y está más o menos cubierta por los bordes de ésta; suele ser de consistencia membranosa, a menudo hialina y posee por lo común dos nervaduras prominentes formando dos carenas. La pálea puede estar atrofiada o, incluso, faltar.
- Flor. La flor es desnuda (carece de cáliz y corola), pero está acompañada generalmente por dos (raramente tres) pequeñas piezas escamosas traslúcidas, denominadas lodículas, que constituyen un resto de perianto. La flor puede ser hermafrodita o unisexual. En muchos géneros los antecios llevan flores hermafroditas, salvo las superiores que son masculinas. En cambio, en muchas Paníceas, hay un antecio inferior masculino y uno superior hermafrodita. Otras veces hay espiguillas masculinas y espiguillas femeninas en la misma planta (el caso de Zea mays, especie diclino monoica) o en plantas distintas (ejemplo, en el subgénero Dioicopoa de Poa, constituido por especies diclino dioicas). Las lodículas parecen ser restos de un perianto trímero y todavía existen 3 lodículas en algunos géneros de Bambúseas y en algunas especies de Stipa. La turgencia de las lodículas determina la apertura del antecio permitiendo la salida al exterior de los estambres y estigmas (momento que se denomina floración o antesis). La morfología de las lodículas tiene importancia sistemática. Asimismo, las flores suelen poseer alguno de estos dos tipos de compresión, importantes taxonómicamente: o están comprimidas lateralmente (de forma que la lemma y la pálea se observen a uno y otro lado de la cara comprimida), o están comprimidas ventralmente (de forma que cada cara comprimida posea a la lemma o a la pálea).
- Androceo. El androceo en las gramíneas es cíclico y en general trímero (es decir, está compuesto por tres estambres, o un múltiplo de tres).
El número más frecuente de estambres es 3, pero en muchas especies de Oríceas y de Bambúseas hay de 6 a 9 (a veces más). En cambio, en Imperata brasiliensis hay un solo estambre y dos en el género Anthoxanthum. Las anteras son basifijas, biloculares y se insertan sobre filamentos delgados y más o menos largos. Los granos de polen son relativamente pequeños, con paredes muy tenues, livianos, tienen un sólo poro germinativo (se dicen monoporados) y están adaptados a ser llevados por el viento desde los estambres hasta los estigmas de otras plantas (la polinización es anemófila).
 Flores en antesis de Holcus mollis, obsérvense los estigmas plumosos y los estambres.
Flores en antesis de Holcus mollis, obsérvense los estigmas plumosos y los estambres. - Gineceo. El gineceo está formado por un ovario globoso, piriforme o fusiforme, generalmente bicarpelar, unilocular, con dos estilos cortos y estigmas plumosos. En algunas Bambúseas el ovario es tricarpelar y lleva 3 estilos. Cuando hay 3 carpelos, la pieza adaxial es fértil (Kircher 1986).[6] En Euchlaena y Zea existe un solo estilo acrescente. Dentro del ovario, súpero, hay un solo óvulo anfítropo o semi-anátropo, subapical o casi basal, de placentación parietal. Algunos pastos tienen óvulos péndulos, átropos. La pared del megasporangio puede ser delgada o gruesa. En muchas especies de gramíneas existen flores cleistógamas en inflorescencias reducidas situadas en las axilas de las hojas inferiores.[2] [3]
- Androceo. El androceo en las gramíneas es cíclico y en general trímero (es decir, está compuesto por tres estambres, o un múltiplo de tres).
Inflorescencia secundaria
Las espiguillas, a su vez, se hallan reunidas o agrupadas en inflorescencias compuestas de tipo racimoso. Las más frecuentes son:
- Panojas laxas, son racimos de espiguillas con pedicelos muy largos. Ejemplo: Avena.
- Panojas densas, son racimos de espiguillas con pedicelos muy cortos. Pueden ser continuas (denominadas espiciformes) o interrumpidas. Ejemplo de la primera es Phalaris, y de la segunda, Dactylis.
- Panojas espateoladas, en las cuales cada racimo terminal o cada pequeña panoja posee una bráctea que lo separa de los otros. Ejemplo, Schizachyrium.
- Racimos espiciformes, con espiguillas muy brevemente pediceladas dispuestas a los dos lados o a un solo lado del raquis. Ejemplo, Paspalum.
- Espiga de espiguillas, con espigas sésiles dispuestas a un solo lado del raquis o espigas unilaterales; en dos series alternándose sobre los lados opuestos del raquis, o espigas dísticas (Lolium), o en varias series sobre el raquis, o espigas cilíndricas (Zea).[2] [3]
Fruto
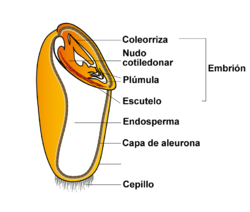 Esquema de un cariopse de gramínea, mostrando el endosperma, el embrión y diversas estructuras embrionales: el escutelo o escudete, la plúmula, la coleorriza, y el nudo cotiledonar. Rodeando a todo el endosperma se aprecia la capa de aleurona y al otro extremo del embrión, el cepillo.
Esquema de un cariopse de gramínea, mostrando el endosperma, el embrión y diversas estructuras embrionales: el escutelo o escudete, la plúmula, la coleorriza, y el nudo cotiledonar. Rodeando a todo el endosperma se aprecia la capa de aleurona y al otro extremo del embrión, el cepillo. Cariopses de arroz (Oryza sativa) en corte longitudinal, los embriones se hallan teñidos de color azul, el endosperma en blanco.
Cariopses de arroz (Oryza sativa) en corte longitudinal, los embriones se hallan teñidos de color azul, el endosperma en blanco.El fruto o grano de las gramíneas es un cariopse, fruto seco indehiscente, con una semilla cuya testa está soldada con el pericarpio formando una envoltura muy delgada. Esta envoltura encierra al embrión y al albumen o endosperma. Este fruto es básicamente una variante del aquenio, aunque se puede encontrar una variedad de frutos en la familia (ver por ejemplo Werker 1997).[7] En algunos géneros como Zizianopsis o Eleusine, el pericarpio no está soldado con la semilla, de modo que el fruto es un aquenio (o un utrículo según otros autores). En algunas Bambúseas el fruto es una nuez o una baya, mientras en el género Sporobolus el pericarpio es mucilaginoso y deja salir a la semilla cuando se embebe en agua. Muchos géneros, como Aristida, Stipa, Piptochaetium, Oryza y casi todas las Paníceas poseen cariopses que se desprenden de la planta envueltos por la lemma y por la pálea. En las Andropogóneas son las glumas las que persisten encerrando al cariopse. En Pennisetum y Cenchrus se desprende toda la espiguilla rodeada de un involucro de cerdas o de espinas. La forma del cariopse varía mucho según los géneros, pudiendo ser casi circular como en Briza, oblongo como en Hordeum, lanceolado como en Poa hasta casi lineal, como en Vulpia. En la parte inferior del cariopse visto dorsalmente, se ve el embrión más o menos elíptico cubierto por el pericarpio transparente. Del otro lado, correspondiente al surco o sutura carpelar, se distingue, también por transparencia, la mácula hilar o hilo (o zona de unión de la semilla con el carpelo), que puede ser punctiforme, como en Poa y en las Paníceas, ovada, como en Briza subaristata, o lineal, como en Hordeum, Vulpia o Festuca.
- Embrión. El embrión de las gramíneas es estructuralmente muy complicado y consta de la plántula unida a su cotiledón laminar, altamente modificado, llamado escudete. El cotiledón es delgado, parenquimatoso, llevando en su parte exterior una capa de células epiteliales que durante la germinación secretan enzimas que hidrolizan las sustancias de reserva localizadas en el endosperma. La planta consta de un nudo cotiledonar, donde se inserta el cotiledón, una yémula cubierta de un capuchón o coleoptilo y una radícula envuelta por otro capuchón o coleorriza. En muchos géneros en la parte externa del nudo cotiledonar hay una escama diminuta, el epiblasto, que para algunos autores constituye un resto de un segundo cotiledón, mientras que otros consideran que se trata de un apéndice de la coleorriza.[2] [3]
Cariología
El tamaño y el número de cromosomas tienen gran importancia en la sistemática de las Gramíneas. Hay dos tipos cromosómicos extremos: el tipo festucoide caracterizado por presentar cromosomas grandes y número básico predominantemente x=7 y el tipo panicoide con cromosomas pequeños y números básicos predominantes x=9 y x=10. El tipo festucoide se encuentra en casi todas las tribus de la subfamilia Festucoideae (sin.: Pooideae), con algunas excepciones. Por ejemplo, la tribu Stipeae de esta subfamilia posee cromosomas pequeños y números básicos x=9, 10, 11, 12, 14, 16 y 17. Las restantes subfamilias de las gramíneas presentan el tipo cromosómico panicoide, con cromosomas pequeños y predominio del número básico x=9 y 10. En las Bambusóideas, Ehrhartóideas y Arundinóideas los cromosomas son pequeños y el número básico es x=12. En la subfamilia Danthonioideae se presentan cromosomas de tamaño intermedio y número básico x=6 y 7. La subfamilia Chloridoideae presenta cromosomas pequeños y varios números básicos, x=7, 8, 9, 10, 11, 12 y 14. Las Panicóideas siempre tienen cromosomas pequeños, con números básicos x=9 o x=10, aunque existen especies con otros números básicos, que varían desde x=4 a x=19.[3]
El genoma de las gramíneas
Las gramíneas son morfológicamente distintas de cualquier otra familia de plantas y, además, son muy diversas en cuanto a morfología y hábito de crecimiento. Las diferentes especies de gramíneas — como se ha descrito en la sección previa — difieren en sus tamaños y números cromosómicos. Asimismo, difieren en el tamaño (o contenido de ADN) de sus genomas. El genoma del arroz, por ejemplo, es más de 11 veces más pequeño que el genoma de la cebada, a pesar de que ambas especies son diploides y aparentan tener la misma complejidad morfológica y fisiológica.[8] El contenido de genes de las diferentes especies de gramíneas, no obstante, no varia tan ampliamente como el contenido de ADN total. El arroz y la cebada, nuevamente, no difieren más que en 2 veces en el número promedio de fragmentos de restricción que hibridan con las mismas sondas.[9] La mayor parte de las diferencias en el tamaño del genoma entre especies de gramíneas se debe a diferencias en el ADN repetitivo. Los genomas más grandes, como los de cebada o trigo, están compuestos en un 75% de ADN repetitivo, mientras que los genomas más pequeños, como el del arroz, solo contienen menos del 50% de ADN altamente repetitivo. Más aún, se ha determinado que buena parte de ese ADN repetitivo está compuesto de retrotransposones insertos entre los genes.[10] [11] Los estudios de mapeo genómico en muchas especies de gramíneas utilizando las mismas sondas de ADN han demostrado que no solo el contenido de genes está muy conservado, sino también el orden de los genes dentro de los cromosomas. La extensa conservación en el contenido de genes y en el orden de los mismos entre el maíz y el sorgo no es inesperada ya que ambas especies "sólo" cuentan con 15 a 20 millones de años de evolución independiente. No obstante, similares observaciones para el arroz y el maíz, las cuales divergieron hace 60 a 80 millones de años, indican que todas las especies de la familia provienen de un mismo antepasado común y que todas ellas conservan un mismo repertorio de genes en el mismo orden aproximado.[12] [13] [14] Los grandes rearreglos genómicos que diferencian entre sí a todas las gramíneas son el resultado de inversiones, translocaciones o duplicaciones cromosómicas que involucran la mayor parte de los brazos cromosómicos.[15] La mayoría, sino todas, las gramíneas son poliploides. Basados en el supuesto que todos los géneros y familias que presentan un número cromosómico básico x=12 son derivados de ancestros que sufrieron duplicaciones cromosómicas durante su evolución y que las subfamilias de gramíneas más primitivas (Anomochlooideae, Pharoideae, y Puelioideae) tienen un número cromosómico básico x=12, se puede deducir que el ancestro de las gramíneas ya era un poliploide. Se sigue, además, que todas las gramíneas que se clasifican como diploides son, en realidad, paleopoliploides (o sea, poliploides antiguos que presentan herencia disómica[16] y cuyos progenitores no pueden ser identificados mediante herramientas citogenéticas o marcadores moleculares).[17] [18] La familia contiene más del 60% de especies, distribuidas en todos los clados, que se clasifican como neopoliploides, o sea que han sufrido un ciclo adicional de duplicación genómica. En estas especies, los genomas duplicados no han divergido mucho del genoma de sus ancestros y su número de cromosomas y comportamiento citológico durante la meiosis son indicativos de la duplicación cromosómica que los ha originado. La mayoría de estos neopoliploides (más del 65%) han derivado de cruzamientos interespecíficos o intergenéricos por lo que se les clasifica como alopoliploides.[17]
Fitoquímica
Las hemicelulosas y los polisacáridos de pectinas de la pared celular primaria de los pastos son muy diferentes de los de las demás espermatofitas, tanto en estructura como en las particularidades de la composición de los xiloglucanos (O'Neill y York 2003).[19] Los polisacáridos son menos ramificados que en todas las demás familias de plantas, si bien esta afirmación está basada en un muestreo todavía escaso de especies. Las poáceas pueden ser cianogenéticas o no. Cuando son cianogenéticas, los compuestos cianogenéticos son derivados de la tirosina. Pueden presentar alcaloides (a veces isoquinolina, pirrolizidina e indol). Raramente puede haber proantocianidinas y cianidinas, en cantidades traza, y sólo en representantes de las subfamilias Panicoideae y Chloridoideae. Los flavonoides se han hallado sólo en algunos géneros, Bouteloua Glyceria y Melica,[20] cuando están presentes son quercetina, o kaempferol junto con quercetina. El ácido elágico y la arbutina no se han encontrado en ningún miembro de la familia. Raramente se encuentran saponinas y sapogeninas, así como también oxalatos libres (por ejemplo en Setaria).
Sistemas reproductivos
Una generalización acerca del modo de reproducción de las gramíneas es que los miembros de esta familia son plantas hermafroditas, que presentan fertilización cruzada (son alógamos) y se polinizan por el viento. Obviamente, una familia con cerca de 10.000 especies cuenta con muchas excepciones a esta regla, las cuales se describen a continuación.
Dioecia
Este tipo de sistema reproductivo, en el cual existen plantas femeninas y plantas masculinas, no es muy frecuente en las gramíneas. Sólo 18 géneros son dioicos o presentan especies dioicas, siendo Poa el más conocido de ellos. De hecho, las especies dioicas de Poa se incluyen en un subgénero separado, Dioicopoa.[21]
Ginodioecia
Este sistema reproductivo describe el hecho de que en las poblaciones naturales de una especie coexisten individuos femeninos e individuos hermafroditas. Esta condición es bastante rara en las gramíneas. Bouteloua chondrosioides y algunas especies del subgénero Andinae de Poa son ginodioicas, si bien Cortaderia es el ejemplo más conspicuo.[22]
Monoecia
En este sistema los sexos están separados espacialmente pero en el mismo individuo, o sea, cada planta presenta inflorescencias femeninas y masculinas. Zea, Humbertochloa, Luziola, Ekmanochloa y Mniochloa son ejemplos de géneros con especies monoicas. Mucho más común entre las gramíneas son las especies andromonoicas, una condición muy común en las Andropogóneas y Paníceas. En las primeras, los dos sexos se presentan en espiguillas diferentes de pares heterógamos. Un par heterógamo de espiguillas consiste usualmente en una espiguilla sesil, con una flor neutra y otra hermafrodita, y una espiguilla pedicelada, con una flor neutra y otra masculina. En las espiguillas bifloras de las Paniceas, en cambio, la flor inferior es usualmente masculina o neutra, y la superior es hermafrodita. Algunos de los géneros que ejemplifican este tipo de sistema son Alloteropsis, Brachiaria, Cenchrus, Echinochloa, Melinis, Oplismenus, Panicum, Setaria, Whiteochloa, y Xyochlaena. Algunas especies dentro de estos géneros pueden tener sólo flores hermafroditas ya que la flor inferior es siempre neutra, raramente ambas flores son hermafroditas. Aparte de las paníceas y las andropogóneas, Arundinelleae es otra tribu con especies andromonoicas. En el resto de la familia, las especies andromonoicas se encuentran muy esporádicamente, como por ejemplo en Arrhenatherum, Hierochloe y Holcus.[22]
Autoincompatibilidad
La gran mayoría de las especies de gramíneas son hermafroditas, no obstante, frecuentemente son incapaces de producir semillas cuando el polen de una planta poliniza sus propios estigmas. Esto se debe a que una gran parte de las especies de la familia presentan autoincompatibilidad, de tipo gametofítica y debido a la acción de dos genes independientes (llamados S y Z) con varios alelos cada uno. Este sistema de autoincompatibilidad ha sido observado en varios géneros de la familia (Festuca, Secale, Lolium, Hordeum, Dactylis, entre muchos otros) y no es perfectamente eficiente. De hecho, de la mayoría de las especies autoincompatibles puede obtenerse una proporción — si bien reducida — de semillas al autofecundar una planta.[22] [23]
Autocompatibilidad
La autopolinización y la autofecundación están muy distribuidas entre las gramíneas. En general, es un mecanismo más común entre las especies anuales que entre las perennes y, decididamente, mucho más frecuente entre las especies colonizadoras. Este mecanismo se ha determinado en aproximadamente 45 géneros de gramíneas, entre los cuales se hallan géneros económicamente muy importantes como Triticum, Oryza, Secale, Avena, Agropyron y Lolium. Una condición de autogamia extrema es la cleistogamia, en la cual se produce la polinización y la fecundación dentro del antecio sin que se produzca la antesis. Este último sistema está distribuido en más de 70 géneros pertenecientes a 20 tribus de gramíneas.[22] [24]
Apomixis
La apomixis se define como la reproducción asexual a través de semillas. En este sistema reproductivo los embriones se desarrollan por mitosis a partir de una oósfera no reducida sin que tenga lugar la fecundación. En otras palabras, cada embrión producido es genéticamente idéntico a la planta madre. En las gramíneas, la apomixis fue descrita por primera vez en 1933 en una especie de Poa. Desde aquel momento se ha identificado este mecanismo en cientos de especies de poáceas, particularmente en las Paníceas y en las Andropogóneas. Algunos de los géneros que presentan especies apomícticas son Apluda, Capillipedium, Heteropogon, Themeda, Sorghum, Bothriochloa, Dichanthium, Cenchrus, Setaria y Paspalum.[22] [25] [26]
Ecología
Asociaciones con otros organismos
Las micorrizas y las micofilas son dos tipos de mutualismos en los cuales se hallan involucrados plantas superiores y hongos. Las micorrizas son mutualismos entre hongos y las raíces de las plantas. Las micofilas, entre tanto, son mutualismos entre hongos endófitos (aquellos que crecen dentro de las plantas) y la parte aérea de las plantas.[27] [28] Por otro lado, las gramíneas también pueden asociarse con diferentes géneros de bacterias de vida libre que fijan el nitrógeno atmosférico.
Micofilas
La presencia de hongos endófitos puede modificar la supervivencia de las plantas de varios modos ya que se pueden producir 3 tipos diferentes de asociaciones entre los simbiontes fúngicos y las plantas, las cuales varían de acuerdo con el grupo taxonómico del hospedante, con las estructuras fúngicas y vegetales involucradas y con las particularidades intrínsecas de la simbiosis (Saikkonen et al. 1998;[29] Gundel et al. 2006).[30] Así, el tipo I tiene como hospedantes a miembros de la familia Juncaceae y varias subfamilias de Poaceae. La colonización es total y el hongo produce sus estructuras sexuales en estromas, suprimiéndose la formación de cariopses en la planta. Toda la población hospedante pierde la capacidad de reproducirse sexualmente. La interacción resulta sumamente agresiva para el hospedante y determina su decaimiento o enfermedad. El tipo II tiene como hospedantes a las Poaceae de la subfamilia Pooideae. No todos los individuos de la población colonizada presentan estromas; en los ejemplares sin síntomas externos el micelio endofítico llega a colonizar las cariopses sin perjudicar la reproducción sexual de la gramínea. De esta manera, el carácter patógeno de la interacción es menor que en el caso anterior. Por último, en el tipo III, que también se da en la subfamilia Pooideae, nunca aparecen estromas en los hospedantes; la colonización del hongo es sistémica, alcanzando a las cariopses por medio de las cuales se propaga dicha asociación. Este tipo de interacción es considerado una simbiosis mutualista debido a que los endófitos benefician a los hospedantes aumentando su crecimiento, biomasa, tasa fotosintética, tolerancia a las heladas y sequías, resistencia a nematodos e insectos, por lo que incrementan la competitividad de sus hospedantes. Asimismo, debido a que producen alcaloides protegen a las plantas del ataque de un amplio rango de animales herbívoros, constituyendo así parte de su sistema de defensa. La presencia de endófitos afecta la palatabilidad de los pastos para los herbívoros y también la palatabilidad de las semillas para los pájaros granívoros, los animales que comen material infectado presentan diversos síntomas de intoxicación (Madej y Clay 1991[31] para las aves (cf.[32] )). El nivel de la infestación por áfidos y la de sus parásitos y parasitoides, y aun el patrón y la tasa de descomposición del pasto muerto, también son afectados por el mutualismo (Omacini et al. 2001[33] para los áfidos; Lemons et al. 2005[34] para la descomposición). Es más, las larvas de moscas del género Phorbia (o Botanophila) viven en el estroma de los hongos endofitos del género Epichloë, y los adultos transmiten los espermatios en una forma análoga a la polinización por insectos de las flores (Bultman 1995[35] ). A su vez, los endófitos se benefician recibiendo el aporte directo de hidratos de carbono producidos por sus hospedantes. De las 232 micofilas conocidas en el mundo, 209 tienen como hospedantes a miembros de todas las subfamilias de Poaceae y representan a los tres tipos de interacción. Las áreas de distribución de esta interacción abarcan tanto zonas frías y templadas como tropicales. En el Hemisferio Norte son muy frecuentes las interacciones tipo I, II y III, mientras que en el Hemisferio Sur prevalecen las de tipo III.[36]
 Claviceps purpurea en una espiga de trigo. Se observan dos espiguillas cuyos granos han sido reemplazados por estructuras sin forma definida: el cuerpo de fructificación del hongo.
Claviceps purpurea en una espiga de trigo. Se observan dos espiguillas cuyos granos han sido reemplazados por estructuras sin forma definida: el cuerpo de fructificación del hongo.Los hongos endofitos de la familia Clavicipitaceae están ampliamente distribuidos entre las gramíneas. Un género de esta familia de hongos, Epichloë, es un endofito restringido a la subfamilia Pooideae (Schardl et al., 1997,[37] Schardl et al., 2004,[38] Craven et al., 2001,[39] Clay y Schardl, 2002[40] y Gentile et al., 2005[41] ) Neotyphodium es el estado imperfecto (o asexual) de Epichloë (Roberts et al., 2004[42] ). Leuchtmann (1992[43] ) discute la distribución y especificidad de huésped de los endofitos de pastos en general (Clay 1990[44] ). Más del 30 % de las especies de Pooideae están involucradas en tales asociaciones, y existe transmisión de estos hongos (la subfamilia Balansiae de las Clavicipitáceas) tanto en forma vertical como horizontal.[45] Las micofilas son una asociación que parece datar de hace unos 40 millones de años y una de sus consecuencias es la producción de alcaloides como, por ejemplo, la lolina (pirrolizidina).[46] Los alcaloides producidos, como la mencionada lolina, son principalmente activos en la defensa de las plantas contra insectos (Schardl et al., 2007[47] ). Otras muchas especies endofitas, aparentemente asintomáticas, pueden crecer conjuntamente en las gramíneas, pero se conoce muy poco sobre sus relaciones. Márquez et al. (2007[48] ), por ejemplo, informaron que Dicanthelium lanuginosum sólo puede crecer en suelos calentados por acción volcánica cuando el hongo endofítico Curvularia, con el que está asociado, se halla infectado con un virus. Esto indica que las relaciones entre las gramíneas y los hongos endófitos pueden ser extremadamente complejas y sus efectos insospechados. Existen listados de especies de hongos endófitos asociados con innumerables especies de gramíneas, como por ejemplo, la provista por Tang y colaboradores en 2007.[49] ) De hecho, sólo en los bambúes hay al menos unas 1.933 especies de hongos descritas.
Micorrizas
Los registros fósiles más antiguos indican que esta asociación tiene unos 400 millones de años, lo que indica la compleja coevolución entre las plantas y sus hongos asociados, que se manifiesta en la amplia distribución del fenómeno (se ha estimado que el 90% de las plantas terrestres están micorrizadas) y en la diversidad de mecanismos morfológicos, fisiológicos y ecológicos implicados. Durante la simbiosis, la planta hospedera recibe nutrientes minerales del suelo tomados por el hongo (principalmente fósforo), mientras que éste obtiene compuestos de carbono derivados de la fotosíntesis.[50] Los hongos formadores de micorrizas arbusculares (HFMA) constituyen micorrizas que colonizan el tejido interno de las raíces de la planta hospedera, donde desarrollan estructuras características de la simbiosis (arbúsculos y vesículas), así como micelio extrarradical, el cuál interacciona con el ecosistema de la rizósfera y es el encargado de la toma de nutrientes del suelo.[51] En este sentido las gramíneas no son la excepción. Buena parte de las especies de esta familia forman micorrizas, lo que favorece y optimiza su adaptación a diversos tipos de ambientes.[52]
Asociación con bacterias fijadoras de nitrógeno
El nitrógeno molecular (N2) es la única reserva de nitrógeno accesible en la biósfera. Prácticamente ilimitada, esta reserva no es directamente utilizada por los vegetales y animales. El nitrógeno es un constituyente esencial de moléculas fundamentales de todos los seres vivos: aminoácidos, proteínas, ácidos nucleicos, vitaminas, entre las más importantes. Para que el nitógeno atmosférico pueda ser asimilado, es necesario que sea reducido. Las gramíneas son capaces de asociarse con bacterias diazotróficas pertenecientes a los géneros Azospirillum, Azotobacter, Azoarcus y Herbaspirillum las cuales realizan la fijación biológica del nitrógeno atmosférico N2). Estas bacterias son organismos de vida libre capaces de fijar nitrógeno desde la rizósfera, o sea, desde el área circundante al sistema radicular de la planta e incluirlo en compuestos (como el amonio) fácilmente disponible y absorbible por las plantas. Además de fijar el nitrógeno atmosférico, las bacterias diazotróficas favorecen el desarrollo del sistema radicular de la planta con la cual conviven, al parecer a través de la producción de reguladores de crecimiento u hormonas. De este modo, favorecen una mayor absorción de nutrientes por parte de la planta. Se han informado incrementos del orden de 5 % hasta 30 % en los rendimientos de gramíneas como caña de azúcar, maíz, arroz, trigo y gramíneas forrajeras como resultado de esta asociación. Estas asociaciones no desarrollan estructuras diferenciadas en las que se alberguen los microorganismos, como ocurre en el caso de las leguminosas y las bacterias del género Rhizobium. En 1998 se ha descrito otro tipo de asociación de diazótrofos en la cual la bacteria (llamada bacteria endófita) se localiza en el interior de la raíz, el tallo y las hojas de la planta. Esta asociación fue descubierta en aislamientos de diazótrofos de plantas forrajeras de Pakistán, en donde se identificó una nueva bacteria fijadora de nitrógeno llamada Azoarcus. Este microorganismo se localiza en las capas externas del córtex; una vez en el interior de la planta se disemina a los tejidos aéreos probablemente por medio de los vasos del xilema.[53] [54]
Distribución
Las gramíneas son una familia cosmopolita que habita desde los desiertos hasta los hábitats de agua dulce o marinos, y todas las elevaciones salvo las más altas del planeta. En el mundo se han desarrollado extensas praderas nativas de gramíneas donde hay sequías periódicas, topografía plana o inclinada, incendios frecuentes, y en algunas ocasiones donde hay pastoreo y bajo ciertas condiciones particulares de suelo. Las comunidades dominadas por los pastos suman el 24 % de la vegetación del planeta, ejemplos son las praderas de Norteamérica, las pampas de Sudamérica, el "veldt" o la sabana en África, y las estepas euroasiáticas (en España, las estepas ibéricas). Por fuera de las praderas herbáceas, los bambúes leñosos desempeñan un papel central en la ecología de los bosques de Asia tropical y templada.
Otros aspectos ecológicos
 Pradera natural de gramíneas. Oglala National Grassland, Estados Unidos.
Pradera natural de gramíneas. Oglala National Grassland, Estados Unidos.Las gramíneas han sido ecológicamente exitosas y se han diversificado extensamente debido a muchas adaptaciones clave. La espiguilla protege a las flores pero, al mismo tiempo, permite la polinización cuando las lodículas abren el antecio. Asimismo, las espiguillas (lemmas con pelos o ganchos) poseen varias adaptaciones para la dispersión del fruto. La versatilidad en los sistemas de apareamiento, incluyendo la autofecundación y la apomixis, permitió a diversas especies de gramíneas ser colonizadoras exitosas de nuevos ambientes. La anatomía de la hoja, que puede ser C3 o C4, permite a estas especies explorar y adaptarse a un amplio rango de hábitats. Los meristemas se hallan ubicados en la base de los entrenudos y en la base de las vainas, protegidos por toda la planta, lo que da como resultado una adaptación al pastoreo y al fuego sin igual entre todas las plantas. El desarrollo de las praderas durante el Mioceno (hace unos 25 a 5 millones de años) puede haber fomentado la evolución de los grandes herbívoros, además de representar una importante fuente de alimento y un estímulo para la evolución del Homo sapiens.
Las gramíneas son anemófilas, es decir que, para efectuar la polinización, el polen es transportado de una planta a otra por medio del viento. Ninguna poácea tiene nectarios, si bien la polinización por insectos se conoce en algunos pastos de bosques, especialmente pequeñas bambusoideas (Soderstrom y Calderón 1971[55] ). La dispersión de las semillas se produce principalmente por animales, y si bien unas pocas gramíneas tienen frutos verdaderos (una excepción es Alvimia, una bambusoidea) puede haber otras estructuras que atraigan a los animales, como los elaiosomas (Davidse 1987[56] ), así como ganchos y agujas mediante los cuales las diásporas se pegan a los animales que pasan (un buen ejemplo es Centotheca). Un gran número de especies es dispersado por el viento, por ejemplo, por tener largos pelos en las aristas, mientras que Spinifex y unos pocos otros géneros son plantas rodantes. Las aristas pueden ayudar tanto en la dispersión por viento como en la dispersión por animales; la microestructura de la superficie de las aristas puede dar como resultado que el cariopse sea directamente "plantado" en el suelo (Elbaum et al. 2007[57] ). Davidse (1987[56] ) remarca un número de taxones con "diásporas reptadoras" ("creeping diaspores").
Los miembros leñosos de los bambúes son conocidos por florecer en forma sincronizada, y muchos son monocárpicos (es decir que luego de florecer y dar la semilla, mueren); esta característica también se encuentra en algunos bambúes herbáceos y, dependiendo de las relaciones de parentesco dentro de la subfamilia, también puede ser un carácter plesiomórfico (ancestral). En plantas que muestran este tipo de reproducción, todos los miembros de un mismo clon florecen simultáneamente, sea donde fuere que hayan sido transportados en todo el globo terrestre, y todas las plantas, después de un período reproductivo que se puede decir retrasado, mueren.
En los tallos huecos de los bambúes muchas veces se junta agua, y vive en ella una fauna distintiva. Las poáceas proveen alimento tanto para los adultos (el polen) como para las larvas (las raíces) de distintas especies de escarabajos de la subfamilia Galerucinae de los crisomélidos.(Jolivet y Hawkeswood 1995[58] ). Las orugas de mariposas de la familia Nymphalidae, en particular las marrones, Satyrinae, y las emparentadas Morphinae, son comunes en los miembros de esta familia (se encuentran en alrededor del 10 % de los censos). Los insectos del taxón Hemiptera-Lygaeidae-Blissinae han sido observados más comúnmente en el clado llamado PACCMAD, menos comúnmente en el clado llamado BEP. Teracrini también se encuentra concentrado en las poáceas (Slater 1976[59] ).
Son comunes en las poáceas los hongos parásitos del orden Uredinales y los de la clase Ustilaginomycetes. Los que atacan a las subfamilias Bambusoideae y Pooideae (incluyendo Stipa y parientes cercanos) son particularmente distintivos (Savile 1979[60] ). Las dos terceras partes de los Ustilaginales (unas 600 especies) se encuentran en Poaceae (Kukkonen y Timonen 1979;[61] Stoll et al. 2003[62] ).
Importancia económica
 Cosecha del trigo (Triticum aestivum), uno de los principales cereales del mundo.
Cosecha del trigo (Triticum aestivum), uno de los principales cereales del mundo. Hojuelas de maíz (Zea mays), uno de los innumerables productos de los cereales.
Hojuelas de maíz (Zea mays), uno de los innumerables productos de los cereales. Vacas pastando sobre una pradera de gramíneas.
Vacas pastando sobre una pradera de gramíneas. Chrysopogon zizanioides, una especie perfumífera. Se observan atados de raíces de vetiveria listos para la venta, Isla de la Reunión.
Chrysopogon zizanioides, una especie perfumífera. Se observan atados de raíces de vetiveria listos para la venta, Isla de la Reunión. Malta utilizada en la elaboración de whisky.
Malta utilizada en la elaboración de whisky. Pleita, artesanía de esparto. Feria de Biodiversidad Agrícola de Bullas, Murcia, España.
Pleita, artesanía de esparto. Feria de Biodiversidad Agrícola de Bullas, Murcia, España. Muchas gramíneas son excelentes fijadoras de dunas. En la imagen se observa una de tales especies, Ammophila arenaria, en Grenen, Dinamarca.
Muchas gramíneas son excelentes fijadoras de dunas. En la imagen se observa una de tales especies, Ammophila arenaria, en Grenen, Dinamarca.La familia de las gramíneas es probablemente la que mayor importancia tiene para la economía humana.[2] De hecho, alrededor del 70% de la superficie cultivable del mundo está sembrada con gramíneas y el 50% de las calorías consumidas por la humanidad proviene de las numerosas especies de gramíneas que son utilizadas directamente en la alimentación, o bien, indirectamente como forrajes para los animales domésticos. En términos de la producción global, los 4 cultivos más importantes son gramíneas: caña de azúcar (Saccharum officinarum), trigo, arroz y maíz. La cebada y el sorgo están entre los primeros 12. Por otro lado, varias especies de gramíneas se utilizan en la industria.
- Plantas alimenticias. Se utiliza generalmente el cariopse o grano directamente como alimento o bien molido en forma de harina. Las especies utilizadas de este modo son los llamados cereales. El hombre ha cultivado cereales por al menos 10.000 años. Desde los comienzos de su domesticación, el trigo (Triticum aestivum), la cebada (Hordeum vulgare), y la avena (Avena sativa) en el Creciente fértil del Próximo Oriente, el sorgo (Sorghum bicolor) y el mijo (Pennisetum americanum) en África, el arroz (Oryza sativa) en el sudeste de Asia, y el maíz (Zea mays) en Meso-América han hecho posible el asentamiento de comunidades humanas y el desarrollo de civilizaciones. El trigo (en especial Triticum aestivum, el denominado trigo pan, una Pooideae), provee la quinta parte de las calorías consumidas por los humanos, y empezó a ser domesticado hace unos 10.000 años. La mayor parte de las formas domésticas son poliploides, y la plasticidad del genoma en conexión con la poliploidía está implicada en el éxito del cultivo de este cereal (Dubcovsky y Dvorak 2007[63] ). El trigo duro (Triticum durum) se utiliza para elaborar fideos o pastas. El maíz (Zea mays, un miembro de Panicoideae) es un cereal con múltiples aplicaciones, desde su consumo directo como "choclo", el uso de su harina en la elaboración de muchos platos regionales e, incluso, bebidas alcohólicas, la utilización industrial de sus granos para la elaboración de aceite, jarabe de fructosa, y muchas otras aplicaciones, a las que se ha agregado últimamente la fabricación de biodiésel. El arroz (Oryza sativa, Ehrhartoideae) es, con toda probabilidad, la especie de mayor importancia global como alimento, dado la enorme cantidad de personas que lo consumen diariamente. La avena (Avena sativa, Pooideae), la cebada (Hordeum vulgare) y el centeno (Secale cereale) son otros tres cereales que se utilizan comúnmente como alimentos. Además de los cereales, algunas gramíneas como Phyllostachys edulis y Sinocalamus beecheyanus se utilizan como hortalizas en Asia.[2]
- Plantas forrajeras. Muchísimas especies de gramíneas son excelentes productoras de pasto para el ganado, tanto en pastizales naturales como en pasturas cultivadas. Así, se cultivan para este objetivo numerosas especies de pastos perennes, tanto en climas templados como en climas tropicales o subtropicales. Las especies forrajeras templadas producen pasto durante el otoño, el invierno y la primavera y las más populares son el raigrás perenne (Lolium perenne), la cebadilla criolla o bromo de los prados (Bromus unioloides), la festuca alta (Festuca arundinacea), el agropiro alargado (Thinopyrum ponticum ), el fleo (Phleum pratense) y el falaris bulboso Phalaris tuberosa). Las gramíneas cultivadas como forrajeras perennes provenientes de climas tropicales o subtropicales son de producción estival y entre ellas se destacan el pasto miel (Paspalum dilatatum), el pasto elefante (Panicum elephantypes), la grama rodes (Chloris gayana), el mijo perla (Pennisetum americanum), el pasto horqueta (Paspalum notatum) y el pasto llorón (Eragrostis curvula). Otras varias especies de forrajeras son anuales, por lo que se las utiliza para producir grandes cantidades de pasto (llamados verdeos) durante una época de producción determinada: el invierno o el verano. Entre las especies para verdeos invernales se hallan la avena (Avena fatua, Avena sativa), el centeno y la cebada forrajera (Hordeum vulgare). Para los verdeos estivales se utiliza el maíz y el sorgo forrajero (Sorghum sudanense). Además hay cereales que se utilizan para alimentar animales, como el mijo (Panicum miliaceum) y el alpiste (Phalaris arundinacea) para alimentar pájaros, o el maíz y el sorgo para alimentar vacas, cerdos y aves de corral.[2] [64]
- Industriales. Los usos industriales de las gramíneas son tan variados como lo es la familia en si misma. Existen especies perfumíferas, cuyos extractos son utilizados en la preparación de una numerosa cantidad de perfumes, tales como el pasto limón (Cymbopogon citratus, del cual se extrae una esencia llamada citronela) y el vetiver (Vetiveria zizanioides). Otras gramíneas, como por ejemplo la cebada cervecera (Hordeum vulgare), son utilizadas para la elaboración de malta, producto indispensable para la fabricación de cerveza, whisky, ginebra, gin y otras bebidas alcohólicas.[65] Otros granos de cereales se utilizan para elaborar bebidas alcohólicas por fermentación, como por ejemplo el sake (o nihonshu) a partir del arroz en Japón. La industria aceitera también utiliza granos de gramíneas (como por ejemplo maíz) para elaborar aceites comestibles. Algunos géneros poseen gran interés en cordelería, cestería y fabricación de calzados tradicionales, como las esparteñas. Tal es el caso del albardín (Lygeum spartum) y, sobre todo, de la atocha (Stipa tenacissima), ambas muy empleadas en España y norte de África para elaborar esparto, materia prima para la confección de todos esos elementos. Asimismo, también se utiliza a Sorghum technicum para fabricar escobas, a Epicampes microura y Aristida pallens para fabricar cepillos y a Stipa tenacissima para elaborar estropajos.[2]
- Gramíneas para césped: numerosas especies de los géneros Poa, Lolium, Festuca, Axonopus, Stenotaphrum y Paspalum se utilizan para formar céspedes en parques y jardines. Agrostis palustris, en particular, se utiliza para formar el "green" de las canchas de golf.[2]
- Gramíneas ornamentales. Hasta no hace muchos años las únicas gramíneas cultivadas en los jardines eran aquellas especies que conforman el césped. En la actualidad muchas gramíneas perennes de gran porte han incrementado su popularidad para ser utilizadas como elementos centrales en el diseño de parques y jardines. Esto no sólo se debe a sus características de longevidad, adaptación, rusticidad y bajo mantenimiento, sino también a sus valores ornamentales o cualidades estéticas particulares relacionados con el movimiento, la transparencia, la iluminación y los espectaculares cambios estacionales que protagonizan. Algunas de las especies que se utilizan como ornamentales son Alopecurus pratensis, Cortaderia selloana, Festuca glauca, Imperata cylindrica, Leymus condensatus, Miscanthus sinensis, Pennisetum setaceum y Phyllostachys aurea, entre muchas otras.[66]
- Malezas: existen numerosas especies de gramíneas que son malezas de diferentes cultivos, varias de ellas muy difíciles de erradicar o controlar, las que ocasionan grandes pérdidas de rendimiento todos los años por competencia con la especie cultivada, como por ejemplo el sorgo de alepo (Sorghum halepense), el gramón (Cynodon dactylon), el capín (Echinochloa crus-galli), el pasto cuaresma (Digitaria sanguinalis) y la braquiaria (Brachiaria extensa).[67] En una de las parábolas de Jesús se utiliza el ejemplo de la cizaña (Lolium temulentum), que es también un lugar común en el habla coloquial.
- Otros usos: las gramíneas también son utilizadas para el control de la erosión y como fijadoras de dunas. Ejemplos de especies empleadas para tal fin son Sporobolus arundinaceus, Panicum urvilleanum, Spartina ciliata, Poa lanuginosa, Ammophila arenaria y Elymus arenarius, entre muchas otras. Ciertos instrumentos musicales son fabricados con cañas de gramíneas, tal es el caso de la quena mediante el empleo de Arthrostylidium harmonicum. Otras especies se han venido utilizando durante siglos como plantas medicinales, en particular como diuréticos, como por ejemplo la grama de los boticarios Elymus repens y Cynodon dactylon. Los bambúes, finalmente, son económicamente importantes en muchas áreas tropicales por sus tallos jóvenes comestibles, por su fibra utilizada para fabricar papel, por su pulpa para el rayon, y sus gruesos tallos para la construcción.
Sistemática
Las comparaciones de las secuencias expresas del ADN y la estructura general de los genomas sugieren que Poaceae difiere mucho más de otras monocotiledóneas, como por ejemplo Allium, de lo que difiere este último de una dicotiledónea como Arabidopsis (Kuhl et al. 2004[68] ). Estas conclusiones acerca de las relaciones de las poáceas con las otras familias de monocotiledóneas y con las dicotiledóneas no son sorprendentes. De hecho, las gramíneas son fácilmente reconocibles e identificables de cualquier otra familia, y su monofilia ha sido sustentada tanto por la morfología como por los análisis moleculares de ADN. Los caracteres fenotípicos que apoyan la monofilia de la familia son la inflorescencia bracteada, el perianto reducido, el tipo de fruto y los caracteres del embrión y de la pared del polen. Las similitudes con las ciperáceas (Cyperaceae) en hábito y en tipo de espiguillas representan una evolución convergente, y no una sinapomorfía. De hecho, Cyperaceae está más emparentado con los juncos (Juncaceae) que con las gramíneas, las cuales pertenecen al núcleo de los Poales.
La importancia económica y ecológica de la familia ha motivado la realización de una importante cantidad de estudios sistemáticos. A principios del siglo XIX, las diferencias entre las espiguillas de las pooideas y las panicoideas llevaron a Robert Brown a dividir a la familia en estos dos grupos básicos. A principios del siglo XX, los caracteres de la epidermis de las hojas y el número de cromosomas llevó a la separación de las chloridoideas de las pooideas. A mediados del siglo XX, la anatomía interna de la hoja (con o sin anatomía de Kranz) y los caracteres del embrión (presencia o ausencia de epiblasto y ciertas características del nudo cotiledonar), llevaron al reconocimiento de 5 a 8 subfamilias. Desde fines del Siglo XX, los estudios filogenéticos basados en varias secuencias génicas (rbcL, ndhF, rpoC2, ITS, "granule bound starch syntase I", y fitocromo B) son congruentes con muchas de las relaciones filogenéticas inferidas previamente a través de caracteres estructurales y fisiológicos. Los estudios moleculares más avanzados (Grass Phylogeny Working Group, GPWG 2001) apoyan el reconocimiento de 12 subfamilias (Clark et al. 1995,[69] Soreng y Davis 1998,[70] Hilu et al. 1999,[71] GPWG 2001[72] ). Los 3 primeros linajes que divergieron, en orden, son:
- Anomochlooideae (nativa de Brasil)
- Pharoideae (nativa de los trópicos del Viejo y Nuevo Mundo)
- Puelioideae (nativa del oeste de África)
Juntos forman sólo unas 25 de las casi 10.000 especies de la familia. El resto de las especies se distribuyen en 2 grandes grupos. El primero, también llamado el clado BEP, agrupa a Bambusoideae sensu stricto, a Ehrharttoideae, y a Pooideae. El segundo, también llamado el clado PACCAD, agrupa a Panicoideae, Arundinoideae sensu stricto, Chloridoideae, Centothecoideae, Aristidoideae, y Danthonioideae. La creación de una nueva subfamilia dentro del clado PACCAD en 2007, Micrairoideae, ha llevado a la redesignación del clado con el acrónimo PACCMAD.[73] El clado PACCMAD está sustentado por un carácter del embrión: un largo entrenudo en el mesocótilo. La anatomía C3 es el estado plesiomórfico (ancestral) en la familia. Todas las especies C4 se encuentran en el clado PACCMAD, aunque muchos subclados C3 se encuentran dentro de él.
Todas las subfamilias reconocidas por el GPWG 2001 son monofiléticas, aunque sólo unas pocas poseen sinapomorfías morfológicas que caractericen a todos sus miembros. Más bien su monofilia es sostenida por grupos de caracteres morfológicos que deben observarse en conjunto. Las relaciones dentro de los grandes clados PACCMAD y BEP son en su mayor parte poco claras todavía, de hecho la posición de Pooideae es poco clara en algunos análisis (Hodkinson et al. 2007,[74] y referencias), sin embargo, Duvall et al. (2007[75] ) encontró buen apoyo para el clado BEP, con un muestreo pequeño.
El cladograma que muestra las relaciones entre estos clados es el siguiente:(GPWG 2001[72] ):
Poaceae clado BEP clado PACCAD Arundinoideae sensu stricto
Posteriormente el Angiosperm Phylogeny Website agregó la subfamilia Micrairoideae (que el GPWG 2001 había dejado sin ubicar), dejando al clado PACCAD como clado PACCMAD, y formó el cladograma siguiente (APWeb 2007):
Poaceae clado BEP clado PACCMAD Arundinoideae sensu stricto
A continuación se brinda una descripción de estas subfamilias y de los géneros más importantes o representativos dentro de cada una.
Anomochlooideae Potzdal
Esta subfamilia fue originalmente incluida en Bambusoideae, pero ahora está claro que sólo poseen un parentesco distante. Estas especies son herbáceas y poseen unas inflorescencias de morfología única que son difíciles de interpretar, y ciertamente no se parecen a las espiguillas características del resto de los pastos. Parecen ser el clado hermano de todo el resto de la familia (Clark et al. 1995,[69] Soreng y Davis 1998[70] ), esto sugiere que la espiguilla característica de los pastos probablemente se originó luego de que Anomochlooideae divergiera. Las plantas de esta subfamilia presentan pseudopecíolo con un pulvínulo apical, la lígula como un mechón de pelos y las ramas de la inflorescencia cimosas. Con respecto a las brácteas de la inflorescencia, presentan 2 a lo largo de cada ramificación y dos más debajo de cada flor. Las anteras son centrifijas (en Anomochloa) o casi basifijas (Streptochaeta), o las flores están dispuestas en forma espiralada a lo largo de ejes racimosos, con varias brácteas debajo de cada flor. Característicamente, la primera hoja de la plántula carece de lámina. El número cromosómico básico es x=11 y 18. Incluye a dos géneros (Anomochloa y Streptochaeta) con 4 especies que habitan selvas y se distribuyen desde Centroamérica hasta el sudeste de Brasil.
- Sinonimia: Anomochloaceae Nakai y Streptochaetaceae Nakai
Pharoideae Clark & Judziewicz
Esta subfamilia, junto con Puelioideae, fue tradicionalmente incluida en Bambusoideae. Presenta espiguillas convencionales que poseen una sola flor en Pharoideae (y muchas flores en Puelioideae). El pasaje de 3 a 2 estigmas ocurrió después de la divergencia de Pharoideae pero antes del origen de Puelioideae.
Pharoideae se caracteriza por sus hojas resupinadas, con las láminas invertidas (con la cara abaxial hacia arriba). Las espiguillas son unifloras y presentan 6 estambres con las anteras centrifijas. El coleoptile presenta lámina. Es una subfamilia que incluye cuatro géneros y unas 12 especies pantropicales, las cuales habitan en las selvas.
- Sinonimia: Pharaceae (Stapf) Herter
Puelioideae Clark, Kobay., Mathews, Spangler & Kellogg
Esta subfamilia, junto con las que vienen más adelante, presenta flores con dos estigmas y espiguillas que se desarticulan por encima de las glumas a la madurez. Presentan un androceo con 6 estambres. Incluye dos géneros (Guaduella y Puelia) los que comprenden aproximadamente 11 especies distribuidas por África tropical.
Bambusoideae Luersson
 Bosque de bambúes, Japón.
Bosque de bambúes, Japón.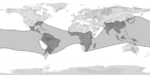 Distribución de las bambusoideas en el mundo.
Distribución de las bambusoideas en el mundo.Las bambusoideas, en sentido restringido, poseen tanto plantas herbáceas como leñosas, y son casi exclusivamente tropicales en su distribución. Las hojas son pseudopecioladas. Presentan 3 lodículas y un androceo con 6 estambres, raramente de 2 a 14. El ovario contiene 2 ó 3 estigmas, raramente 1. La primera hoja de las plántulas no presenta lámina. Los números cromosómicos básicos son x=7 y x=9 a 12. Incluye 126 géneros con aproximadamente 970 a 1.200 especies tropicales a templadas.
La diversificación dentro de los bambúes ocurrió hace 30 a 40 millones de años. Los bambúes leñosos forman un grupo monofilético hermano del clado que contiene a las especies herbáceas. Los bambúes leñosos, con sus tallos de hasta 40 metros de altura, ciertamente no se parecen al césped. La floración en muchas de estas especies también es inusual, ya que ocurre en ciclos de hasta 120 años. Aun cuando los tallos individuales viven por sólo una o unas pocas décadas, alguna forma de "reloj" hace que los tallos florezcan todos al mismo tiempo en todo el rango de distribución de la especie, causando a veces bruscos cambios ecológicos, como los que se asocian a las ratadas. Algunos géneros importantes de bambúes leñosos son Bambusa (120 especies), Chusquea (100 especies), Arundinaria (50 especies), Sasa (50 especies), y Phyllostachys (45 especies).
- Sinonimia: Bambusaceae Burnett, Parianaceae Nakai.
Ehrhartoideae Link
Presentan espiguillas con glumas muy reducidas y el androceo con 6 estambres (raramente 1). Los números cromosómicos básicos de esta subfamilia son x=10 y x=15. Comprende de 105 a 120 especies, entre las que se incluyen a los miembros de las tribus Ehrharteae del Hemisferio Sur, así como al cosmopolita Oryzeae. El último es acuático o de tierras húmedas. El representante más conocido de la tribu Oryzeae es el arroz asiático Oryza sativa, uno de los cultivos más importantes del mundo. También se cultiva otra de las 22 especies de Oryza, O. glaberrima en el oeste de África. En Estados Unidos cobra importancia otra especie de la tribu, Zizania aquatica, el arroz silvestre norteamericano. Guo y Ge (2005[76] ) presentaron una filogenia de la tribu Oryzeae.
- Sinonimia: Oryzaceae Burnett
Pooideae Bentham
Las poóideas presentan oligosacáridos derivados de la fructosa en el tallo, las ramificaciones principales de la inflorescencia son dísticas, la lemma usualmente consta de 5 nervios. Presentan cromosomas usualmente largos y el número cromosómico básico es x=7, más raramente x=2, 4, 5 ó 6. Tiene una distribución predominantemente templada, especialmente en el Hemisferio Norte. La subfamilia incluye unas 3.300 especies. Entre los géneros sobresalientes se incluyen importantes cereales, como el trigo, la cebada y la avena, y también al centeno (Secale cereale), a los pastos utilizados para césped (como Poa, con 500 especies), para heno (Festuca, 450 especies), para pasturas (como Phleum, Dactylis), y algunas malas hierbas (como Agrostis, con 220 especies, y Poa). Otros géneros importantes de esta subfamilia son Stipa (300 especies), Calamagrostis (270 especies), Bromus (150 especies), y Elymus (150 especies).
- Sinonimia: Aegilopaceae Martynov, Agrostidaceae Burnett, Alopecuraceae Martynov, Avenaceae Martynov, Festucaceae Pfeiffer, Hordeaceae Burnett, Melicaceae Martynov, Nardaceae Martynov, Phalaridaceae Burnett, Stipaceae Burnett, Triticaceae Hochst.
Chloridoideae Beilschmied
 Inflorescencias de Eragrostis tef, una cloridoidea.
Inflorescencias de Eragrostis tef, una cloridoidea.Los integrantes de las cloridoideas presentan espiguillas que se desarticulan por encima de las glumas y poseen pelos bicelulares distintivos en la epidermis de las hojas. No obstante, este último carácter puede ser una sinapomorfía de sólo un subgrupo del clado. Todo el clado salvo dos especies muestra fotosíntesis por la vía del C4. Los números cromosómicos básicos prevalecientes en la subfamilia son x=9 y x=19, aunque existen géneros con x=7 y 8. La subfamilia se desarrolla mejor en regiones tropicales áridas y semiáridas, donde se supone que la fotosíntesis C4 es ventajosa. Los centros de distribución ubicados en África y Australia sugieren un origen en el Hemisferio Norte. Algunos géneros importantes son Eragrostis (350 especies), Muhlenbergia (160 especies), Sporobolus (160 especies), Chloris (55 especies), Spartina (15 especies), y Eustachys (10 especies), los primeros 3 géneros mencionados aparentemente son polifiléticos.
- Sinonimia: Chloridaceae (Berchtold & J. Presl) Herter, Eragrostidaceae (Stapf) Herter, Pappophoraceae (Kunth) Herter, Spartinaceae Burnett, Sporobolaceae Herter
Panicoideae Link
 Distintos colores de granos en espigas de maíz (Zea mays, una panicóidea).
Distintos colores de granos en espigas de maíz (Zea mays, una panicóidea). Inflorescencias de caña de azúcar (Saccharum officinarum , una panicóidea).
Inflorescencias de caña de azúcar (Saccharum officinarum , una panicóidea). Arundo donax, una arundinóidea.
Arundo donax, una arundinóidea. Cortaderia selloana, una dantonóidea.
Cortaderia selloana, una dantonóidea.Las panicóideas han sido reconocidas taxonómicamente desde hace mucho tiempo, debido a sus espiguillas características. Las cañas son usualmente sólidas, las espiguillas se hallan comprimidas dorsalmente, no presentan raquilla y son bifloras. La desarticulación de la espiguilla a la madurez se produce por debajo de las glumas. El tipo de fisiología de la fotosíntesis prevaleciente es la C4. Los gránulos de almidón en el endosperma son simples. Los números cromosómicos básicos más típicos son x=5, 9 y 10, aunque también se hallan especies con x=7, 12 y 14. La subfamilia es principalmente tropical y contiene dos grandes tribus, Andropogoneae y Paniceae, junto con un número de grupos pequeños. Andropogoneae son relativamente fáciles de reconocer porque sus espiguillas suelen venir de a pares y agrupadas en una inflorescencia lineal. Paniceae no es tan homogénea como Andropogoneae. La subfamilia comprende 206 géneros y 3.245 especies. Entre los géneros más importantes se incluyen Panicum (470 especies, polifilético), Paspalum (330 especies), Andropogon (100 especies), Setaria (100 especies), Sorghum (20 especies), y Zea (4 especies). El sorgo y el maíz son dos cultivos de gran importancia económica y ambos se incluyen en esta subfamilia.
- Sinonimia: Andropogonaceae Martynov, Arundinellaceae (Stapf) Herter, Panicaceae Voight, Saccharaceae Martynov, Zeaceae A. Kern
Centothecoideae Soderstrom
Las centotecoideas son una subfamilia pobremente estudiada. Está constituida por unas 30 especies distribuidas en 11 géneros que habitan selvas templadas cálidas a tropicales. Su característica más distintiva es la presencia de estilo en la flor y de epiblasto en el embrión. El número cromosómico básico más frecuente es x=12, aunque también hay géneros con x=11.
Arundinoideae Burmeister
Con 14 géneros y entre 20 a 38 especies, las arundinoideas son una subfamilia cuya exacta delimitación todavía no es clara. Son gramíneas hidrofíticas a xerofíticas que habitan regiones templadas a tropicales. Sus números cromosómicos básicos son x=6, 9 y 12. Arundo (con 3 especies, Arundo donax es la caña de Castilla) y Phragmites (2 especies) son los géneros más conocidos de esta subfamilia.
- Sinonimia: Arundinaceae (Dumortier) Hochst.
Micrairoideae Pilger
Esta subfamilia monofilética perteneciente al clado PACCMAD ha sida reinstalada y circunscripta nuevamente en el año 2007. Los miembros de la misma presentan estomas con células subsidiarias en forma de domo, las lígulas con mechones de pelos, embriones pequeños, fotosíntesis C4, granos de almidón simples en el endosperma. Comprende 8 géneros y unas 170 especies en su mayoría tropicales. Algunos de los géneros (como por ejemplo Eriachne) no estaban asignados a ninguna familia hasta hace poco tiempo y otros estaban incluidos en otras subfamilias (ejemplo Isachne en las Panicoideas).[73] Incluye a los géneros Isachne (100 especies), Eriachne (35 especies) y Micraira (8 especies).
Aristidoideae Caro
Las aristoideas incluyen 3 géneros y de 300 a 385 especies de regiones templadas cálidas, con aristas con una columna basal y fotosíntesis del tipo C4. Los números cromosómicos básicos son x=11 y x=12. Comprende al gran género Aristida (230 a 330 especies) y Stipagrostis (50 especies).
Danthonioideae Barker & Linder
Danthonioideae es una subfamilia bastante distribuida en todo el globo, especialmente en el Hemisferio Sur. Presentan profilos bilobados y las sinérgidas del saco embrionario de tipo haustorial. Los números cromosómicos básicos son x=6, 7 y 9. Comprende 19 géneros y unas 270 especies. Entre los géneros con mayor número de especies se hallan Danthonia (100 especies) y Rytidosperma (90 especies). Cortaderia selloana es la popular "cortadera", en inglés "pampa grass", una gramínea ornamental que se incluye dentro de esta subfamilia.
Taxonomía
- Introducción teórica en Taxonomía
Entre los caracteres diagnósticos de la familia se hallan varios de los órganos ya descriptos: las hojas que tienen vainas largas y abiertas y las lígulas en la unión entre la vaina y la lámina, los tallos redondos y usualmente huecos en los entrenudos, y las inflorescencias cuya unidad básica es la espiguilla. Las flores individuales son pequeñas, con perianto inconspicuo y con un gineceo que usualmente tiene dos estigmas plumosos y un único óvulo. En el fruto seco, de tipo aquenio (cariopse), el embrión —relativamente grande— ocupa una posición lateral y se ubica junto al tegumento de la semilla.
Son particularmente útiles para la identificación de los géneros los caracteres de la espiguilla, como el tamaño, el plano de compresión, la presencia o ausencia de glumas, el número de flores por espiguilla, la presencia de flores estériles o incompletas, el número de venas en las glumas y las glumelas, la presencia o ausencia de aristas, y la forma de las inflorescencias secundarias.
Lista de géneros
El listado de todos los géneros conocidos de gramíneas, ordenados alfabéticamente, se provee en el anexo denominado géneros de Poaceae. En muchos casos se listan sinónimos. Los enlaces a sinónimos llevan al nombre de género preferido.
Sinonimia
La familia o alguna de sus subfamilias poseen los siguientes sinónimos:
Aegilopaceae, Agrostidaceae, Alopecuraceae, Andropogonaceae, Anomochloaceae, Arundinaceae, Arundinellaceae, Avenaceae, Bambusaceae, Chloridaceae, Eragrostidaceae, Festucaceae, Hordeaceae, Lepturaceae, Melicaceae, Miliaceae, Nardaceae, Oryzaceae, Panicaceae, Pappophoraceae, Parianaceae, Phalaridaceae, Pharaceae, Saccharaceae, Spartinaceae, Sporobolaceae, Stipaceae, Streptochaetaceae, Triticaceae, Zeaceae.[77]
Evolución
Las gramíneas y todos sus parientes extintos (lo que en paleontología se llama el "stem group" Poaceae) datan de hace unos 89 millones de años, el grupo principal divergió hace unos 83 millones de años (Janssen y Bremer 2004,[78] Bremer 2002;[79] fechas en Wikström et al. 2001[80] ) Con respecto a los pastos más "típicos" (los clados BEP y PACCMAD), los fósiles de espiguillas asignados a ellos se conocen en el límite entre el Paleoceno y el Eoceno, hace unos 55 millones de años (Crepet y Feldman 1991[81] ), y esta estimación está a grandes rasgos en línea con una estimación de la edad de una duplicación del genoma de las gramíneas (hace unos 70-50 millones de años, Blanc y Wolfe 2004;[82] Schlueter et al. 2004;[83] Paterson et al. 2004[84] ). Sin embargo, Poinar (2004[85] ) sugirió que Programinis burmitis del Cretácico temprano (hace unos 100-110 millones de años) es del tipo bambusoideo temprano. Si bien este fósil tiene un número de caracteres vegetativos que son comunes entre las poáceas, su identidad necesita confirmación.
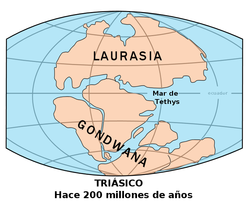 El grupo basal de las gramíneas se habría originado y diversificado en Gondwana antes del aislamiento del subcontinente indio.
El grupo basal de las gramíneas se habría originado y diversificado en Gondwana antes del aislamiento del subcontinente indio.Los tejidos vegetales silicificados (fitolitos) preservados en heces fosilizadas (coprolitos) de dinosaurios del Cretáceo tardío halladas en la India indican que por lo menos 5 taxones pertenecientes a subclados de gramíneas extinguidas estaban presentes en el subcontinente indio durante ese período geológico (hace unos 71-65 millones de años). Esta diversidad taxonómica sugiere que el grupo basal (crown-group) de las gramíneas se habría diversificado y distribuido en Gondwana antes de que la India quedara geográficamente aislada (Prasad et al. 2005[86] ).
Más allá de estos fósiles, la fotosíntesis C4 parece haber estado presente en las gramíneas del Mioceno temprano a medio, tanto en las Grandes Planicies de Norteamérica como en África, hace unos 25-12,5 millones de años. Quizás este tipo de fotosíntesis estuvo inicialmente asociada a cambios adaptativos en respuesta a una disminución en la concentración de CO2 en la atmósfera, si bien la gran expansión de este mecanismo fisiológico ocurrió hace sólo unos 9-4 millones de años. Si este evento estuvo además favorecido por los incrementos en la temperatura, la disminución de las precipitaciones, el aumento de los vientos y el concomitante incremento de incendios, que habrían removido a los árboles de algunos hábitats en ese período, aún no está claro (Edwards y Still 2007,[87] y Tipple y Pagani 2007,[88] para los orígenes tempranos de la fotosíntesis C4 y su subsecuente desarrollo; Jacobs et al. 1999[89] para la paleoecología de Poaceae; Fox y Koch 2004;[90] Osborne y Beerling 2006[91] ). Los detalles de los mecanismos de la fotosíntesis C4 y las morfologías asociadas con ella son muy diversos y presentan una considerable variación, particularmente en el caso de la subfamilia de las panicóideas. De hecho, la fotosíntesis C4 aparentemente se originó y evolucionó independientemente hasta 8 veces en esta subfamilia (Giussani et al. 2001[92] ). Asimismo, este mecanismo se originó de modo independiente en otras subfamilias, como Micrairoideae, Aristidoideae y Chloridoideae (Kellogg, 2000[93] ). Independientemente de su mayor eficiencia fotosintética, las gramíneas C4 presentan menor contenido de nitrógeno, mayor cantidad de fibras de esclerénquima y pueden ser menos palatables que las gramíneas C3 (Caswell et al. 1973[94] ). A pesar de estas características, existió una radiación de mamíferos pastadores en el Mioceno (Thomasson y Voorhies 1990[95] ) que pudo estar asociada con la ampliación de las praderas y sabanas, dominadas por pastos. Sin embargo, cuando las especies de gramíneas de pradera se expandieron en Nebraska en el Mioceno temprano —hace unos 23 millones de años— los ungulados hipsodontos ya existían para entonces (Strömberg 2004[96] ).
Las gramíneas han desarrollado un gran número de características fisiológicas que les han permitido conquistar diversos hábitats donde prevalecen condiciones subóptimas para el crecimiento de las plantas. Una característica ecológica quizás relacionada de un gran número de especies distribuidas en diferentes subfamilias es la capacidad de acumular betaínas de glicina y otros compuestos que se hallan asociados con la adaptación de las plantas al crecimiento en condiciones salinas (Rhodes y Hanson 1993[97] ). Por otro lado, el núcleo de Pooideae almacena carbohidratos como fructanos, los que se hallan en mucha menor concentración en las restantes especies de la familia. Esta característica está asociada con la adaptación de tales especies a condiciones de estrés hídrico (sequías) y de bajas temperaturas (heladas, Hendry 1993[98] ). Finalmente, otro mecanismo fisiológico singular de las poáceas es que aparentemente son la única familia de angiospermas que adquiere iones por quelación de iones férricos con sideróforos que son absorbidos por las raíces (Schmidt 2003[99] ).
La inflorescencia de las gramíneas es una estructura nueva en el repertorio reproductivo de las plantas con flores. Es intrincada desde el punto de vista de la biología del desarrollo, tiene una importancia central en la agronomía y, finalmente, es una verdadera intriga evolutiva. Su arquitectura controla el tipo de polinización y la producción de semillas, por lo que es una diana importantísima tanto para la selección natural como para el mejoramiento genético y la biotecnología. No es sorprendente, entonces, que la diversidad de estructuras que presentan las espiguillas y espigas de las gramíneas estén controladas por genes que afectan el desarrollo que no están presentes en ninguna otra familia de plantas. Estos genes se han originado luego de extensas duplicaciones del genoma y la posterior diversificación funcional de los mismos.[100] [101] [102] [103] [104] [105] Algunos de los genes que afectan el desarrollo floral y que se ha demostrado que se hallan duplicados en las gramíneas son, por ejemplo, las familias de genes API, AG y SEP AP1/FUL y LOFSEP.
La pálea (y aparentemente también la lemma) quizás sea derivada del cáliz, y las lodículas quizás sean derivadas de la corola (Ambrose et al. 2000[106] ). Por otro lado un estudio de morfología comparativa sugiere que la lemma es una bráctea y que la pálea representa dos tépalos connados del verticilo más externo (Whipple y Schmidt 2006[107] ). Teniendo en cuenta las relaciones de parentesco cercano entre Ecdeicoleaceae y Joinvilleaceae recientemente encontradas por Marchant y Briggs (2007[108] ) y la probabilidad de que las flores de Anomochloa sean sui generis, la morfología floral de Streptochaeta puede ser plesiomórfica (ancestral) en la familia. Es interesante que las flores de Ecdeicolea también son notablemente monosimétricas, con los dos tépalos adaxiales del verticilo externo más largos y aquillados, y si bien esto no es relevante en forma directa, una diferenciación comparable en el verticilo externo del perianto ocurre en Xyridaceae, todos estos probablemente sean paralelismos. Una interpretación más común de la pálea es que es de naturaleza profilar/bracteolar, las monocotiledóneas comúnmente tienen profilos bicarenados, sin embargo, parece que las bracteolas tuvieron que reaparecer en Poaceae, ya que los clados más emparentados pero externos a la familia ("outgroups" en análisis cladístico) no las poseen. Las lodículas parecen estar involucradas en la apertura de las flores estaminadas y las perfectas, mientras que pueden estar ausentes en las flores pistiladas (Sajo et al. 2007[109] ).
Referencias citadas
- ↑ Judd, W. S., Campbell, C. S. Kellogg, E. A. Stevens, P.F. Donoghue, M. J. (2002), «Poaceae» Plant systematics: a phylogenetic approach, Sinauer Axxoc, 287-292. ISBN 0-87893-403-0.
- ↑ a b c d e f g h i j Parodi, L.R. 1987. Gramíneas. En: Enciclopedia Argentina de Agricultura y Jardinería. Tomo I. Descripción de plantas cultivadas. Editorial ACME S.A.C.I., Buenos Aires, PP: 108-182
- ↑ a b c d e f Cabrera, A. L., Cámara Hernández, J., Caro, J.; Covas, G.; Fabris, H., Hunziker, J., Nicora, E., Rugolo, Z., Sánchez, E., Torres, M. (1970), «Gramineae, parte general.» Flora de la Provincia de Buenos Aires: Gramíneas., Colección Científica del INTA. Tomo IV, parte II., 1-18.
- ↑ Se denomina "artejo" a cada uno de los segmentos claramente delimitados en que se divide un órgano articulado, en este caso se refiere a cada uno de los segmentos en los que se divide una espiga de gramínea. ver glosario.
- ↑ Carenada es un término que se utiliza en botánica para referirse a cualquier órgano de la planta que presenta la forma de una quilla o carena. Ref.: Parodi, L.R. 1987. Gramíneas. En: Enciclopedia Argentina de Agricultura y Jardinería. Tomo I. Segundo Volumen. Descripción de plantas cultivadas. Editorial ACME S.A.C.I., Buenos Aires, PP: 1114.
- ↑ Kircher, P. (1986). Untersuchungen zur Blüten- und Infloreszenmorphologie, Embryologie und Systematik der Restionaceen im vergleich mit Gramineen und verwandten Familien. [Dissertationes botanicae 94.]. Berlin: J. Cramer. ISBN.
- ↑ Werker, E. (1997). Seed Anatomy [Handbuch der Pflanzenanatomie 10, 3.]. Berlin: Borntraeger. ISBN.
- ↑ Arumuganathan, K. & E.D. Earle. 1991. Nuclear DNA content of some important plant species. Plant Mol. Biol. Rep. 9: 208-218.Resumen en inglés
- ↑ Kurata, N., G. Moore, Y. Nagamura, T. Foote, M. Yano, Y. Minobe, and M.D. Gale. 1994. Conservation of genome structure between rice and wheat. BioTechnology 12: 276-278 resumen en inglés
- ↑ Flavell, R.B., M.D. Bennett, J.B. Smith, and D.B. Smith. 1974. Genome size and proportion of repeated nucleotide sequence DNA in plants. Biochem. Genet. 12: 257-269
- ↑ SanMiguel, P., A. Tikhonov, Y.-K. Jin, N. Motchoulskaia, D. Zakharov, A. Melake-Berhan, P.S. Springer, K.J. Edwards, Z. Avramova, and J.L. Bennetzen. 1996. Nested retrotransposons in the intergenic regions of the maize genome. Science 274: 765-768 resumen en inglés
- ↑ Clark, L.G., W. Zhang, and J.F. Wendel. 1995. A phylogeny of the grass family (Poaceae) based on ndhF sequence data. Syst. Bot. 20: 436-460
- ↑ Bennetzen, J.L. and M. Freeling. 1993. Grasses as a single genetic system: Genome composition, collinearity and compatibility. Trends Genet. 9: 259-261
- ↑ Bennetzen, J.L. & Freeling, M. 1997. The Unified Grass Genome: Synergy in Synteny Genome Res.7: 301 - 306.Artículo en inglés
- ↑ Moore, G., T. Foote, T. Helentjaris, K. Devos, N. Kurata, & M. Gale. 1995. Was there a single ancestral grass chromosome? Trends Genet.
- ↑ Herencia disómica es el tipo de herencia que presentan los organismos diploides, los que llevan sólo dos juegos de cromosomas. Aplicado a un poliploide indica que, desde el punto de vista genético y pese a estar constituido por varios juegos de cromosomas, ese organismo se comporta como un diploide.
- ↑ a b Levy, A. & Moshe Feldman. 2002. The Impact of Polyploidy on Grass Genome Evolution Plant Physiol. 130: 1587-1593 Artículo en inglés
- ↑ La paleopoliploidía se puede detectar sólo a través de herramientas bioinformáticas bastante sofisticadas que revelan la similitud y colinearidad entre genes que divergieron decenas de millones de años atrás.
- ↑ O'Neill, M. A.; York, W. S. (2003). «The composition and structure of plant primary cell walls.». En Rose, J. K. C.. The Plant Cell Wall. Oxford: Blackwell. pp. 1-54. ISBN.
- ↑ Herrera, Y. & Chairez, I. 2005. Flavonoids of the genus Bouteloua (Poaceae) from Mexico. Núm.20, pp.17-29, ISSN 1405-2768; México. Artículo en inglés
- ↑ Connor, H.E. 1979. Breeding systems in the grasses: a survey. New Zealand Journal of Botany 17:547-74. Resumen en inglés
- ↑ a b c d e Connor, H.E. 1979. Breeding systems in the grasses: a survey. New Zealand Journal of Botany 17:547-74.
- ↑ Baumann, U., Juttner, J., Bian, X. & P. Langridge. 2000. Self-incompatibility in the Grasses. Annals of Botany 85 (Supplement A): 203-209. Artículo en inglés
- ↑ Campbell, C., J. Quinn, G. Cheplick, & T J Bell. 1983. Cleistogamy in Grasses. Annual Review of Ecology and Systematics Vol. 14: 411-441.
- ↑ Savidan, Y. 2000. Apomixis: Genetics and Breeding. Plant Breeding Reviews, Volume 18, Jules Janick (ed). John Wiley & Sons, Inc. ISBN 0-471-35567-4. Artículo en inglés
- ↑ Kellogg, E. 1990. Variation and Species Limits in Agamospermous Grasses. Systematic Botany 15: 112-123Resumen en inglés
- ↑ Lewis, D.H. (1987). «Evolutionary aspects of mutualistic associations between fungi and photosynthetic organisms». En A. D. M. Rayner, C. M. Brasier & D. Moore. Microbiology of the Phyllosphere. (Evolutionary biology of the fungi edición). Cambridge University Press, Cambridge. ISBN.
- ↑ May, T. (1997). «Fungi». En Scott, G.A.M, Entwisle, T.J., May, T.W. & Stevens, G.N.. A Conservation Overview of Australian Non-marine Lichens, Bryophytes, Algae and Fungi.. Environment Australia, Canberra. ISBN 0-642-21399-2.
- ↑ Saikkonen, K.; Faeth, S. H., Helander, M., y Sullivan, T. J. (1998). «Fungal endophytes: A continuum of interactions with host plants.». Ann. Rev. Ecol. Syst. (29): pp. 319-343.
- ↑ Gundel, P. E.; Maseda, P. H., Vila-Aiub, M. M., Chersa, G. M., y Benech-Arnold, R. (2006). «Effects of Neotyphodium fungi on Lolium multiflorum seed germination in relation to water availablity.». Ann. Bot. (97): pp. 571-577.
- ↑ Madej, C. W.; Clay, K. (1991). «Avian seed preference and weight loss experiments: The effects of fungal endophyte-infected tall fescue seeds.». Oecologia (88): pp. 296-302.
- ↑ Jamieson IG and HS. Easton Does a toxic fungal endophyte of tall fescue affect reproduction of takahe on offshore islands? DOC SCIENCE INTERNAL SERIES 89
- ↑ Omacini, M.; Chaneton, E. J., Ghersa, C. M., y Müller, C. B. (2001). «Symbiotic fungal endophytes control insect host-parasite interaction webs.». Nature (409): pp. 79-81.
- ↑ Lemons, A.; Clay, K., & Rudgers, J. A. (2005). «Connecting plant-microbial interactions above and below ground: A fungal endophyte affects decomposition.». Oecologia (145): pp. 595-604.
- ↑ Bultman, T. L. (1995). «Mutualistic and parasitic interactions between Phorbia flies and Epichloë: Convergence between a fungus and entomophilous angiosperms.». Canadian J. Bot. (73(suppl. 1)): pp. S1343-1348.
- ↑ Clay, K.; Schardl, C. (2002). «Evolutionary origins and ecological consequences of endophyte symbiosis with grasses.». American Naturalist (160): pp. S99-S127.
- ↑ Schardl, C. L.; Leuchtmann, A., Chung, K. R., Penny, D., y Siegel, M. R. (1997). «Coevolution by common descent of fungal symbionts (Epichloë spp.) and grass hosts.». Mol. Biol. Evol. (14): pp. 133-143.
- ↑ Schardl, C. L.; Leuchtmann, A., y Spiering, M. J. (2004). «Symbioses of grasses with seedborne fungal endophytes.». Ann. Rev. Plant Biol. (55): pp. 315-340.
- ↑ Craven, K. D.; Hsiau, P. T. W., Leuchtmann, A., Hollin, W., & Schardl, C. L. (2001). «Multigene phylogeny of Epichloë species, fungal symbionts of grasses.». Ann. Missouri Bot. Gard. (88): pp. 14-34.
- ↑ Clay, K.; Schardl, C. (2002). «Evolutionary origins and ecological consequences of endophyte symbiosis with grasses.». American Naturalist (160): pp. S99-S127.
- ↑ Gentile, A.; Rossi, M. S., Cabral, D., Craven, K. D., y Schardl, C. L. (2005). «Origin, divergence and phylogeny of Epichloë endophytes of native Argentine grasses.». Mol. Phyl. Evol. (35): pp. 196-208.
- ↑ Roberts, C.; West, C., & Spiers, D. (2004). Neotyphodium in Cool-Season Grasses. (Blackwell edición).
- ↑ Leuchtmann, A. (1992). «Systematics, distribution, and host specificty of grass endophytes.». Natural Toxins (1): pp. 150-162.
- ↑ Clay (1990). «Fungal endophytes of grasses.». Ann. Review Ecol. Syst. (21): pp. 275-297.
- ↑ Clay, K. (1986). «Grass endophytes.». En Fokkema, N. J., y van den Heuvel, J.. Microbiology of the Phyllosphere. (Cambridge Universty Press edición). Cambridge. pp. 188-204. ISBN.
- ↑ Schardl, C. L.; Leuchtmann, A., y Spiering, M. J. (2004). «Symbioses of grasses with seedborne fungal endophytes.». Ann. Rev. Plant Biol. (55): pp. 315-340.
- ↑ Schardl, C. L.; Grossman, R. B., Nagabhyru, P., Faulkner, J. R., y Mallik, U. P. (2007). «Loline alkaloids: Currencies of mutualism.». Phytochem. (68): pp. 980-996.
- ↑ Márquez, L. M.; Redman, R. S., Rodriguez, R. J., y Roossinck, M. J. (2007). «The virus in a fungus in a plant: Three-way symbiosis required for thermal tolerance.». Science (315): pp. 513-515.
- ↑ Tang, A. M. C.; Shekoy, B. D., y Hyde, K. D. (2007). «Fungal diversity.». En Hodkinson, T. R., y Parnell. J. A. N.. Reconstructing the Tree of Life: Taxonomy and Systematics of Species Rich Taxa [Systematics Association Special Volume Series 72.] (CRC Press edición). Boca Raton, FLA.. pp. 227-249.
- ↑ Simon, L., Bousquet, R., Levesque, C., Lalonde, M. 1993. Origin and Diversification of Endomycorrhizal Fungi and Coincidence with Vascular Land Plants. Nature 363: 67-69.
- ↑ Hodge, A. 2002. Microbial Ecology of the Arbuscular Mycorrhiza. FEMS Microbiol Ecol.32:91-96.
- ↑ Trappe, J. M. 1987. Phylogenetic and ecological aspects of mycotrophy in the Angiosperms from an evolutionary standpoint.Pp. 5-25. In: G. R. Safir (Ed.) Ecophysiology of VA mycorrhizal plants. CRC Press, Boca Raton.
- ↑ Reinhold-Hurek, B., & Hurek, T., 1998. Life in grasses: diazotrophic endophytes. Trends Microbiol. 139:139-143.
- ↑ Baca, B.E., Soto Urzúa, L. & Pardo Ruiz, M.E. 2000. Fijación biológica de nitrógeno. Elementos 38: 43-49. Artículo en pdf
- ↑ Soderstrom, T. R.; Calderön, C. E. (1971). «Insect pollination in tropical rain forest grasses.». Biotropica (3): pp. 1-16.
- ↑ a b Davidse, G. (1987). «Fruit dispersal in the Poaceae.». En Soderstrom, T. R., Hilu, K. W., Campbell, C. S., y Barkworth, M. E.. Grass Systematics and Evolution. (Smithsonian Institiution Press edición). Washington, D.C.. pp. 143-155. ISBN.
- ↑ Elbaum, R.; Zaltzman, L., Burgert, I., y Fratzl, P. (2007). «The role of wheat awns in the seed dispersal unit.». Science (316): pp. 884-886.
- ↑ Jolivet, P. H.; Hawkeswood, T. J. (1995). «Fungal diversity.». Host-Plants of the Chrysomelidae of the World. (Backhuys edición). Leiden. pp. 227-249.
- ↑ Slater, J. A.; Redman, R. S., Rodriguez, R. J., y Roossinck, M. J. (1976). «Monocots and chinch bugs: A study of host plant relationships in the Lygaeid subfamily Blissinae (Hemiptera: Lygaeidae).». Biotropica (8): pp. 143-165.
- ↑ Savile, D. B. O. (1979). «Fungi as aids in higher plant classification.». Bot. Rev. (45): pp. 377-503.
- ↑ Kukkonen, I.; Timonen, T. (1979). «Species of Ustilaginales, especially of the genus Anthracoidea, as tools in plant taxonomy.». Symb. Bot. Upsalienses (22): pp. 166-176.
- ↑ Stoll, M.; Begerow, D., Piepenbring, M., y Oberwinkler, F. (2003). «Coevolution of "smut fungi" (Ustilaginales) and grasses (Poaceae).». Palm. Hortus Francofurtensis (7): pp. 135.
- ↑ Dubcovsky, J.; Dvorak, J. (2007). «Genome plasticity a key factor in the success of polyploid wheat under domestication.». Science (316): pp. 1862-1866.
- ↑ Langer, R.H.M. 1984. Especies y variedades de gramíneas forrajeras. En: Las pasturas y sus plantas, Capítulo 3, pag.: 75-96. Editorial Hemisferio Sur, Montevideo, Uruguay.
- ↑ Kent, Norman Leslie [1983] Technology of cereals: An introduction for students of food science and agriculture. Pergamon Press Ltd, Oxford.
- ↑ Puglia, M.L. (mayo 2003). «Gramíneas ornamentales». Jardín (Edición especial Nº7): pp. 1-130. ISSN 0329-322.
- ↑ Listado de malezas pertenecientes a las gramíneas. [1]
- ↑ Kuhl, J. C.; Cheung, C., Yuan, Q., Martin, W., Zewdie, Y., McCallum, J., Catanach, A., Rutherford, P., Sink, K. C., Jenderek, M., Prince, J. P., Town, C. D., y Harvey, M. J. (2004). «A unique set of 11,008 onion Expressed Sequence Tags reveals expressed sequence and genomic differences between the monocot orders Asparagales and Poales.». Plant Cell (16): pp. 114-125.
- ↑ a b Clark, L. G.; Zhang, W., Wendel, J. F. (1995). «A phylogeny of the grass family (Poaceae) based on ndhF sequence data.». Syst. Bot. (20): pp. 436-460.
- ↑ a b Soreng, R. J.; Davis, J. I. (1998). «Phylogenetics and character evolution in the grass family (Poaceae): simultaneous analysis of morphological and chloroplast DNA restriction site character sets.». Bot. Rev. (64): pp. 1-85.
- ↑ Hilu, K. W.; Alice, L. A., y Liang, H. (1999). «Phylogeny of Poaceae inferred from matK sequences.». Ann. Missouri Bot. Gard. (86): pp. 835-851.
- ↑ a b Grass Phylogeny Working Group (2001). «Phylogeny and subfamilial classification of the grasses (Poaceae).». Ann. Missouri Bot. Gard. (88): pp. 373-457.
- ↑ a b Sánchen-Ken, J. Gabriel; Clark, Lynn G.; Kellogg, Elizabeth A.; Kay, Elma E. 2007. Reinstatement and Emendation of Subfamily Micrairoideae (Poaceae). Systematic Botany 32, Number 1, 71-80.Resumen en inglés
- ↑ Hodkinson, T. R.; Savolainen, V., Jacobs, S. W. L., Bouchenak-Khelladi, Y., Kinney, M. S. & Salamin, N. (2007). «Supersizing: Progress in documenting and understanding grass species richness.». En Hodkinson, T. R., y Parnell. J. A. N.. Reconstructing the Tree of Life: Taxonomy and Systematics of Species Rich Taxa [Systematics Association Special Volume Series 72.] (CRC Press edición). Boca Raton, FLA. pp. 275-295. ISBN.
- ↑ Duvall, M. R.; Leseberg, C. H., y Grennan, C. P. (2007). «Comparison of whole chloroplast genomes in grasses (Poaceae); evolutionary insights related to Coix lacryma-jobi, Microcalamus convallarioides, Puelia olyriformis and Joinvillea plicata.». Plant Biology and Botany 2007. Program and Abstract Book. Chicago. pp. 284. ISBN.
- ↑ Guo, Y.-L.; Ge, S. (2005). «Molecular phylogeny of Oryzeae (Poaceae) based on DNA sequences from chloroplast, mitochondrial, and nuclear genomes.». American J. Bot. (92): pp. 1548-1558.
- ↑ Cornell University. Poaceae. http://biopl-a-181.plantbio.cornell.edu/. Consultado el 04-4-2008.
- ↑ Janssen, T.; Bremer, K. (2004). «The age of major monocot groups inferred from 800+ rbcL sequences.». Bot. J. Linnean Soc. (146): pp. 385-398.
- ↑ Bremer, K. (2002). «Gondwanan evolution of the grass alliance of families (Poales).». Evolution (56): pp. 1374-1387. http://www.bioone.org/perlserv/?request=get-abstract&doi=10.1554%2F0014-3820(2002)056%5B1374%3AGEOTGA%5D2.0.CO%3B2&ct=1. Consultado el 04-11-2007.
- ↑ Wikström, N.; Savolainen, V., y Chase, M. W. (2001). «Evolution of the angiosperms: Calibrating the family tree.». Proc. Roy. Soc. London B (268): pp. 2211-2220. http://links.jstor.org/sici?sici=0962-8452(20011107)268%3A1482%3C2211%3AEOTACT%3E2.0.CO%3B2-A. Consultado el 04-11-2007.
- ↑ Crepet, W. L.; Feldman, G. D. (1991). «The earliest remains of grasses in the fossil record.». American J. Bot. (78): pp. 1010-1014.
- ↑ Blanc, G.; Wolfe, K. H. (2004). «Widespread paleopolyploidy in model plant species inferred from age distributions of duplicate genes.». Plant Cell (16): pp. 1667-1678. http://www.plantcell.org/cgi/content/abstract/16/7/1667. Consultado el 04-11-2007.
- ↑ Schlueter, J. A.; Dixon, P., Granger, C., Grant, D., Clark, L., Doyle, J. J., y Schoenmaker, R. C. (2004). «Mining EST databases to resolve evolutionary events in major crop species.». Genome (47): pp. 868-876.
- ↑ Paterson, A.; Bowers, J. E., y Chapman, B. A. (2004). «Ancient polyploidization predating divergence of the cereals, and its consequences for comparative genomics.». Proc. National Acad. Sci. U.S.A. (101): pp. 9903-9908.
- ↑ Poinar, G. O. Jr. (2004). «Programinis burmitis gen. et sp. nov., and P. laminatus sp. nov., early Cretaceous grass-like monocots in Burmese amber.». Australian Syst. Bot. (17): pp. 497-504.
- ↑ Prasad, V.; Strömberg, C. A. E., Alimohammadian, H., y Sahni, H. (2005). «Dinosaur coprolites and the early evolution of grasses and grazers.». Science (310): pp. 1177-1180.
- ↑ Edwards, E. J.; Still, C. J. (2007). «Phylogeny and the ecological distribution of C4 grasses.». Plant Biology and Botany 2007. Program and Abstract Book.. Chicago. pp. 183.
- ↑ Tipple, B. J.; Pagani, M. (2007). «The early origins of terrestrial C4 photosynthesis.». Ann. Rev. Earth Planet. Sci. (35): pp. 435-461.
- ↑ Jacobs, B. F.; Kingston, J. D., y Jacobs, B. L. (1999). «The origin of grass-dominated ecosystems.». Ann. Missouri Bot. Gard. (8686): pp. 590-643.
- ↑ Fox, D. L.; Koch, P. L. (2004). «Carbon and oxygen isotopic variability in Neogene paleosol carbonates: Constraints on the evolution of C4-grasslands of the Great Plains, USA.». Palaeogeog. Palaeoclim. Palaeoecol. (207): pp. 305-329.
- ↑ Osborne, C. P.; Beerling, D. J. (2006). «Nature's green revolution: The remarkable evolutionary rise of C4 plants.». Phil. Trans. Roy. Soc. London B (361): pp. 173-194.
- ↑ Giussani, L. M.; Cota-Sánchez, J. H., Zuloaga, F. O., y Kellogg, E. A. (2001). «A molecular phylogeny of the grass subfamily Panicoideae (Poaceae) shows multiple origins of C4 photosynthesis.». American J. Bot. (88): pp. 1993-2012.
- ↑ Kellogg, E. A. (2000). «The grasses: A case study in macroevolution.». Ann. Rev. Ecol. Syst. (31): pp. 217-38.
- ↑ Caswell, H.; Reed, F., Stephenson, S. N., y Werner, P. A. (1973). «Photosynthetic pathways and selective herbivory: A hypothesis.». American Naturalist (107): pp. 465-480.
- ↑ Thomasson, J. R.; Voorhies, M. R. (1990). «Grasslands and grazers.». En Briggs, D. E. G., y Crowther, P. R.. Palaeobiology: A Synthesis. (Blackwell edición). Oxford. pp. 84-87. ISBN.
- ↑ Strömberg, C. A. E. (2004). «Using phyolith assemblages to reconstruct the origin and spread of grass-dominated habitats in the Great Plains of North America during the late Eocene to early Miocene.». Palaeogeogr. Palaeoclim. Palaeoecol. (207): pp. 239-275.
- ↑ Rhodes, D.; Hanson, A. D. (1993). «Quaternary ammonium and tertiary sulfonium compounds in higher plants.». Ann. Rev. Plant Physiol. Plant Mol. Biol. (44): pp. 357-384.
- ↑ Hendry, G. A. F. (1993). «Evolutionary origins and natural functions of fructans - a climatological, biogeographic and mechanistic appraisal.». New Phytol. (123): pp. 3-14.
- ↑ Schmidt, W. (2003). «Iron solutions: Acquisition strategies and signaling pathways in plants.». Trends Plant Sci. (8): pp. 188-193.
- ↑ Malcomber, S. T.; Kellogg, E. A. (2005). «SEPALLATA gene diversification: Brave new whorls.». Trends Plant Sci. (10): pp. 427-435.
- ↑ Schlueter, J. A.; Dixon, P., Granger, C., Grant, D., Clark, L., Doyle, J. J., y Schoenmaker, R. C. (2004). «Mining EST databases to resolve evolutionary events in major crop species.». Genome (47): pp. 868-876.
- ↑ Paterson, A.; Bowers, J. E., y Chapman, B. A. (2004). «Ancient polyploidization predating divergence of the cereals, and its consequences for comparative genomics.». Proc. National Acad. Sci. U.S.A. (101): pp. 9903-9908.
- ↑ Preston, J. C.; Kellogg, E. A. (2006). «Reconstructing the evolutionary history of paralogous APETALA1/FRUITFULL-like genes in grasses (Poaceae).». Genetics (174): pp. 421-437.
- ↑ Malcomber, S. T.; Preston, J. C., Reinheimer, R., Kossuth, J., y Kellogg, E. A. (2006). «Developmental gene evolution and the origin of grass inflorescence diversity.». Adv. Bot. Res. (44): pp. 425-481.
- ↑ Zanis, M. J. (2007). «Grass spikelet genetics and duplicate gene comparisons.». Int. J. Plant Sci. (168): pp. 93-110.
- ↑ Ambrose, B. A.; Lerner, D. R., Ciceri, P., Padilla, C. M., Yanofsky, M. F., y Schmidt, R. J. (2000). «Molecular and genetic analyses of the Silky1 gene reveal conservation in floral organ specification between eudicots and monocots.». Molecular Cell (5): pp. 569-579.
- ↑ Whipple, C. J.; Schmidt, R. J. (2006). «Genetics of grass flower development.». Adv. Bot. Res. (44): pp. 385-424.
- ↑ Marchant, A. D.; Briggs, B. G. (2007). «Ecdeiocoleaceae and Joinvilleaceae, sisters to Poaceae (Poales): Evidence from rbcL and matK data.». Telopea (11): pp. 437-450.
- ↑ Sajo, M. G.; Longhi-Wagner, H., & Rudall, P. J. (2007). «Floral development and embryology in the early-divergent grass Pharus.». Int. J. Plant Sci. (168): pp. 181-191.
Bibliografía
- Stevens, P. F. (2001 en adelante). «Poaceae» (en inglés). Angiosperm Phylogeny Website, versión 8, junio del 2007. Última actualización de la sección: 11-03-2007. Consultado el 04-11-2007.
- Watson, L.; Dallwitz, M. J.. «Poaceae» (en inglés). The families of flowering plants: descriptions, illustrations, identification, and information retrieval. Version: 1st June 2007.. Consultado el 04-11-2007.
- Judd, W. S.; Campbell, C. S. Kellogg, E. A. Stevens, P.F. Donoghue, M. J. (2002). «Poaceae». Plant systematics: a phylogenetic approach (Sinauer Axxoc edición). USA. pp. 287-292. ISBN 0-87893-403-0.
- Cabrera, A. L.; Cámara Hernández, J., Caro, J.; Covas, G.; Fabris, H., Hunziker, J., Nicora, E., Rugolo, Z., Sánchez, E., Torres, M. (1970). «Gramineae, parte general.». Flora de la Provincia de Buenos Aires: Gramíneas. (Colección Científica del INTA. Tomo IV, parte II. edición). Buenos Aires, Argentina.. pp. 1-18.
- Parodi, L.R. (1987). Gramíneas. (En: Enciclopedia Argentina de Agricultura y Jardinería. Tomo I. Descripción de plantas cultivadas. edición). Editorial ACME S.A.C.I., Buenos Aires,.. pp. 108-182.
Lecturas recomendadas
- Watson, L.; Dallwitz, M. J.. «The Grass Genera of the World» (en inglés). Delta-Intkey. Consultado el 04-11-2007. (no posee descripción de la familia, sólo de los géneros).
Enlaces externos
 Wikimedia Commons alberga contenido multimedia sobre Poaceae. Commons
Wikimedia Commons alberga contenido multimedia sobre Poaceae. Commons Wikiespecies tiene un artículo sobre Poaceae. Wikispecies
Wikiespecies tiene un artículo sobre Poaceae. Wikispecies
- Anomochlooideae Potztal
























Wikimedia foundation. 2010.