- Migración de las aves
-
La migración de las aves se refiere a los viajes estacionales regulares realizados por muchas especies de aves. Además de la migración, las aves realizan otros movimientos en respuesta a cambios en la disponibilidad de alimentos, de hábitat o climáticos, que suelen ser irregulares o sólo en una dirección y reciben diversas denominaciones como nomadismo, invasiones, dispersiones o irrupciones. La migración está marcada por su estacionalidad anual.[1] En contraste, las aves que no son migratorias son llamadas aves residentes.
 Una bandada de gansos Branta leucopsis (barnacla de cara blanca) durante la migración de otoño.
Una bandada de gansos Branta leucopsis (barnacla de cara blanca) durante la migración de otoño.
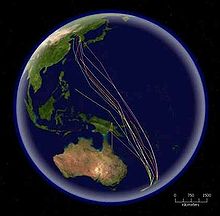
 Algunas rutas migratorias, código de colores: celeste, Oenanthe oenanthe; rojo, Sterna paradisaea; verde, Falco amurensis; azul, Puffinus tenuirostris; amarillo, Philomachus pugnax; rosado, Buteo swainsoni.
Algunas rutas migratorias, código de colores: celeste, Oenanthe oenanthe; rojo, Sterna paradisaea; verde, Falco amurensis; azul, Puffinus tenuirostris; amarillo, Philomachus pugnax; rosado, Buteo swainsoni.Patrones generales
Muchas aves terrestres migran largas distancias. Los patrones más comunes involucran el vuelo al norte para reproducirse en los veranos en áreas templadas o árticas y el retorno a las áreas de invernada en regiones más cálidas del sur.
La ventaja primaria de la migración es energética. Los días más largos del verano en el norte proveen mayores oportunidades a las aves en reproducción de alimentar a sus polluelos. La extensión de las horas del día les permite a las aves diurnas producir nidadas más grandes que las especies emparentadas no migratorias que permanecen en los trópicos todo el año. A medida que los días se acortan en otoño, las aves regresan a las regiones cálidas donde el suministro de alimento disponible varía poco con la estación.
Estas ventajas son superiores a los riesgos del elevado estrés, el costo energético, y otros riesgos de la migración. La predación puede ser incrementada durante la migración; Falco eleonorae (halcón de Eleonora) que cría en las islas del Mediterráneo tiene una temporada de cría muy retrasada, coordinada con el paso otoñal de los pájaros que migran al sur de los que alimenta a sus polluelos. Una estrategia similar es adoptada por el murciélago Nyctalus lasiopterus, que se alimenta de pájaros migratorios.[2] [3] [4] Las mayores concentraciones de aves migrantes en sitios de parada transitoria también los hacen susceptibles de parásitos y patógenos, requiriendo una respuesta inmune más elevada.[5]
Dentro de una especie en concreto, no todas las poblaciones tienen por que ser migratorias, lo que es llamado migración parcial. La migración parcial es muy común en los continentes del sur; en Australia, el 44% de las especies de aves no paseriformes y el 32% de las paseriformes son migradoras parciales[6] En algunas especies la población de latitudes más altas tiende a ser migratoria e invernará frecuentemente en latitudes más bajas que las latitudes en las que las otras poblaciones de la misma especie son sedentarias y tiene por lo tanto ya ocupado ese hábitat apropiado para la invernación, y esto se denomina "migración a saltos de rana".[7] Dentro de una población, también puede existir un patrón diferente de cronología y migración basada en grupos de edades y sexo. Sólo las hembras de Fringilla coelebs (pinzón vulgar) en Escandinavia migran y los machos se quedan como residentes. Esto dio lugar al nombre específico coelebs, que significa soltero.
La mayoría de las migraciones comienzan con las aves despegando en un gran frente. En algunos casos la migración involucra cinturones estrechos de migración que son establecidos como rutas tradicionales denominadas rutas de vuelo migratorio. Éstas siguen típicamente cadenas montañosas y líneas costeras, y pueden tomar ventaja de brisas y otros patrones de viento o evitar barreras geográficas como amplias masas de agua abierta. Las rutas específicas pueden estar genéticamente programadas o aprendidas en grados variados. Las rutas tomadas en un sentido y en el de regreso son a menudo diferentes.[5]
Muchas de las aves más grandes vuelan en bandadas. El volar así las ayuda a reducir el consumo de energía. Muchas grandes aves vuelan en formación de V y los ahorros individuales de energía se han estimado en un monto de 12-20%.[8] [9] Los correlimos Calidris canutus (correlimos gordo) y Calidris alpina (correlimos común) fueron seguidos por estudio de radar encontrándose que volaban 5 km por hora más rápido en bandadas que cuando volaban en solitario.[5]
La altitud a la cual las aves vuelan durante las migraciones varía. Una expedición al Monte Everest encontró esqueletos de Anas acuta (ánade rabudo o pato pescuecilargo) y Limosa limosa (becasina o aguja colinegra) a 5000 m sobre el glaciar Khumbu.[10] Gansos Anser indicus han sido vistos volando sobre los picos más altos de los Himalayas sobre 8000m, incluso cuando había cerca pasos más bajos de 3000m.[11] Las aves marinas vuelan bajo sobre el agua pero ganan altura cuando cruzan sobre tierra y un patrón inverso se ve en las aves terrestres.[12] [13] Sin embargo la mayoría de las migraciones de aves ocurre en el rango de 150 m a 600 m. Los registros de colisión de aves en los Estados Unidos muestran que la mayoría de los impactos ocurren a alturas inferiores a 600 m y casi ninguno por encima de 1800 m.[14]
La mayoría de las especies de pingüinos hacen migraciones regulares nadando. Estas rutas pueden cubrir más de 1000 km Dendragapus obscurus (gallo de las Rocosas) realiza migraciones altitudinales caminando en su mayor parte. Los emúes en Australia han sido observados realizando caminatas de largas distancias durante las sequías.[5]
Visión histórica
Las primeras observaciones registradas de la migración de las aves son de hace 3000 años, mencionadas por Hesíodo, Homero, Heródoto, Aristóteles y otros. La Biblia también menciona migraciones, como en el Libro de Job (39:26), donde se hace la interrogante :
¿Es por tu inteligencia que se cubre de plumas el halcón y despliega sus alas hacia el sur?El autor de Jeremías (8:7) escribió: :
Hasta la cigüeña, en el cielo, conoce sus estaciones; la tórtola, la golondrina y la grulla tienen en cuenta el tiempo de sus migraciones.Aristóteles menciona que las grullas viajan de las estepas escitas a los pantanos de las cabeceras del Nilo. Plinio el Viejo en su Naturalis Historia repite las observaciones de Aristóteles. Por otra parte Aristóteles sugirió que las golondrinas y otras aves hibernaban. Esta creencia persistió incluso hasta 1878, cuando Elliott Coues hizo una lista de no menos de 182 trabajos que trataban sobre la hibernación de las golondrinas. No fue sino hasta principios del siglo XIX en que fue aceptada la migración como causa de la desaparición durante el invierno de las aves en los climas del norte.[15]
El descubrimiento de cigüeñas blancas en Alemania heridas con flechas africanas proveyeron pistas sobre la migración. Uno de los más antiguos especímenes flechados fue encontrado cerca de la aldea alemana de Klütz en el estado de Mecklenburg-Vorpommern.
Migración a larga distancia
 Catharus ustulatus, tordo de espalda olivada.
Catharus ustulatus, tordo de espalda olivada. Anas acuta, ánade rabudo o pato pescuecilargo.
Anas acuta, ánade rabudo o pato pescuecilargo.La imagen típica de la migración son las de las aves terrestres norteñas como las golondrinas y aves rapaces haciendo largos vuelos hacia los trópicos. Muchos patos, gansos y cisnes que se reproducen en el norte son también migrantes de largas distancias, sin embargo necesitarían moverse al sur sólo lo suficiente para escapar de las aguas que se congelan en sus zonas árticas de cría. La mayoría de las especies de Anatidae del Holártico permanecen en el hemisferio norte, pero en países con climas más templados. Por ejemplo, Anser brachyrhynchus (ganso de pico corto) migra desde Islandia a Gran Bretaña y países vecinos. Las rutas migratorias y las zonas de invernada son tradicionales y aprendidas por los jóvenes durante su primera migración junto a sus padres. Algunos patos, como Anas querquedula (cerceta carretota), se mueven completamente o parcialmente a los trópicos.
Las mismas consideraciones acerca de las barreras y contorneos que se aplican a las aves terrestres migrantes a largas distancias, se aplican a las aves acuáticas, pero a la inversa: una gran área de tierra sin acuatorios que ofrezcan un sitio de alimentación es una barrera para un ave acuática. El mar abierto también es una barrera para un ave que se alimenta en las aguas costeras. Se observan contorneos para evitar esas barreras: por ejemplo, Branta bernicla (ganso de collar) que migra de la Península de Taimir al Mar de Frisia (Holanda, Alemania y Dinamarca) viaja por la vía de la costa del Mar Blanco y el Mar Báltico en vez de atravesar directamente el Océano Ártico y el norte de Escandinavia.
 Limosa lapponica, becasina o aguja colinegra.
Limosa lapponica, becasina o aguja colinegra.Una situación similar ocurre con las aves limícolas (Charadriiformes). Muchas especies como Calidris alpina (correlimos común) y Calidris mauri (correlimos de Alaska) hacen largos recorridos desde sus áreas de cría en el Ártico hasta lugares más cálidos del mismo hemisferio, pero otras como Calidris pusilla (correlimos semipalmeado) viajan enormes distancias hacia los trópicos. Como los grandes y poderosos patos y gansos (Anseriformes), las limícolas son grandes voladoras. Esto significa que las aves que invernan en regiones templadas tienen la capacidad de realizar movimientos adicionales cortos en la eventualidad de que se produzca un tiempo particularmente inclemente.
Para algunas especies limícolas, el éxito de la migración depende de que tengan disponibilidad de recursos alimenticios clave en los puntos de parada a lo largo de la ruta migratoria. Esto le da a las migradoras una oportunidad de reabastecerse para el próximo tramo del viaje. Algunos ejemplos de lugares importantes de parada de migratorias son Bahía de Fundy y Bahía de Delaware.
Algunos especímenes de Limosa lapponica (becasina o aguja colipinta) tienen el récord del vuelo sin paradas más largo registrado para una ave migratoria, vuelan 11.000 km desde Alaska hasta sus áreas de estación no reproductiva en Nueva Zelanda.[16] [17] Antes de la migración, el 55 por ciento del peso corporal es grasa acumulada para dar energía a este viaje ininterrumpido.
 Sterna paradisaea, charrán ártico.
Sterna paradisaea, charrán ártico.La migración de las aves marinas es similar en patrón a la de Charadriiformes y Anseriformes. Algunas, como Cepphus grylle (arao de alas blancas) y algunas gaviotas son bastante sedentarias, otras, como la mayoría de los charranes y alcas que crían en zonas templadas del hemisferio norte, se trasladan distancias variables al sur durante el invierno. La mayor distancia de migración entre todas las aves la realiza Sterna paradisaea (charrán ártico) y permanece a la luz del día más que ninguna otra ave, al trasladarse de sus áreas de cría en el Ártico a la zona antártica durante la estación no reproductiva. Un charrán ártico, anillado (marcado con una anilla identificativa) cuando era un pollo en las Islas Farne, mar afuera de las costas orientales británicas, alcanzó Melbourne, Australia, en sólo tres meses después de emplumar; un viaje marítimo de 22.000 km. Unas pocas aves marinas, como Oceanites oceanicus (pamperito de Wilson) y Puffinus gravis (pardela capirotada), crían en el hemisferio sur y migran al norte durante el invierno austral. Las aves marinas tienen la ventaja adicional de poder alimentarse durante la migración sobre aguas abiertas.
Las especies más pelágicas, principalmente Procellariiformes, son grandes vagabundas, y los albatros de los océanos australes pueden circunvolar el globo durante la estación no reproductiva. Las aves Procellariiformes se dispersan ampliamente sobre grandes áreas de océano abierto, pero se congregan cuando el alimento resulta disponible. Muchas están también entre las migrantes a mayores distancias; Puffinus griseus (pardela o pamperito oscuro) que anidan en las Islas Malvinas migran 14.000 km entre la colonia de cría y el norte del Océano Atlántico costa afuera de Noruega. Algunos Puffinus puffinus (pardela pichoneta) hacen este mismo viaje a la inversa. Como son aves muy longevas, pueden sumar enormes distancias recorridas, que en un espécimen fueron calculadas en unos 8 millones de km durante su vida comprobada de más de 50 años.
 Un buitre leonado (Gyps fulvus) planeando.
Un buitre leonado (Gyps fulvus) planeando.Algunas aves grandes de amplias alas dependen de las columnas de aire caliente ascendente para permitirles planear. Esas incluyen muchas aves rapaces como buitres, águilas y gavilanes y también las cigüeñas. Estas aves migran de día. Las aves migratorias de estos grupos tienen gran dificultad para cruzar grandes masas de agua, dado que las columnas termales sólo se forman sobre la tierra, y estas aves no pueden mantener el vuelo activo por largas distancias. El Mediterráneo y otros mares son por lo tanto grandes obstáculos para las aves planeadoras, que son forzadas a cruzar por los puntos más estrechos. Cantidad masivas de grandes rapaces y cigüeñas pasan a través de áreas como Gibraltar, Falsterbo y el Bósforo en tiempos de migración. Las especies más comunes, como Pernis apivorus (halcón abejero), se pueden contar por cientos de miles en otoño. Otras barreras, como las cadenas montañosas, también pueden causar grandes concentraciones, particularmente de migrantes diurnas grandes. Este es un factor notable en el cuello de botella migratorio de América Central.
 Archilochus colubris, colibrí de garganta roja.
Archilochus colubris, colibrí de garganta roja.Muchas de las aves insectívoras más pequeñas, incluyendo las currucas, colibríes y papamoscas, migran largas distancias por lo común de noche. Se posan durante a la mañana y pueden alimentarse durante algunos días antes de continuar la migración. Las aves son llamadas "de paso" en las regiones donde aparecen de forma transitoria en cortas estadías durante el viaje migratorio.[18]
Al migrar durante la noche, las migrantes nocturnas minimizan el riesgo de los depredadores, y evitan el recalentamiento que podría resultar de la energía consumida durante el vuelo en distancias tan largas. Esto también les permite alimentarse durante el día para reponer energía para la noche.[15] Un costo de la migración nocturna es la pérdida de sueño. Las migrantes deben ser capaces de tener un sueño de calidad alterada durante el vuelo para compensar esta pérdida.[19]
Migración a corta distancia
 Bombycilla cedrorum, ampelis o picotero del cedro.
Bombycilla cedrorum, ampelis o picotero del cedro.Muchas de las migrantes de larga distancia de la sección previa son efectivamente programadas genéticamente para responder a la longitud cambiante del día. Sin embargo muchas especies se mueven distancias más cortas, pero lo hacen sólo en respuesta a condiciones climáticas difíciles.
Así que las que se reproducen en montañas y páramos, como Tichodroma muraria (treparriscos) y Cinclus cinclus (mirlo acuático), pueden con sólo moverse altitudinalmente escapar de las tierras altas frías. Otras especies como Falco columbarius (esmerejón) y Alauda arvensis (alondra común) se mueven algo más, hacia la costa o a una región más sureña. Especies como Fringilla coelebs (pinzón vulgar) no son migratorias en Gran Bretaña, pero se moverán al sur o a Irlanda si el tiempo es muy frío.
Las migrantes paseriformes de corta distancia tienen dos orígenes evolutivos. Las que tienen parientes que migran largas distancias en la misma familia, como Phylloscopus collybita (mosquitero común), son especies originalmente del hemisferio sur que han acortado progresivamente su viaje de regreso hasta permanecer en el hemisferio norte.
Las especies que en su familia no tienen parientes de largas migraciones, como en Bombycilla, se mueven solo en respuesta al tiempo invernal, más que por ampliar sus oportunidades reproductivas.
 Halcyon senegalensis, martín pescador arbóreo.
Halcyon senegalensis, martín pescador arbóreo.En los trópicos hay muy poca variación en la duración de las horas de luz a lo largo del año, y siempre es suficientemente cálido como para un adecuado suministro de alimentos. Aparte de los movimientos estacionales de las especies invernantes del hemisferio norte, la mayoría de las especies se mueven distancias variables dependiendo de la pluviosidad.
Muchas regiones tropicales tienen estaciones lluviosas y secas, los mozones de la India son quizá el ejemplo más conocido. Un ejemplo de ave cuya distribución está asociada a la pluviosidad es el martín pescador arbóreo Halcyon senegalensis (alción senegalés) del oeste de África.
Existen unas pocas especies, son notables los cucos, que son verdaderos migrantes de largas distancias dentro de los trópicos. Un ejemplo es Cuculus poliocephalus (cuco o cuclillo menor), que cría en la India y pasa la estación no reproductiva en África.
En las altas montañas, como en las cordilleras de los Himalayas y los Andes, existen también movimientos estacionales altitudinales en muchas especies, y otras pueden realizar migraciones de considerable distancia. Ficedula subrubra (papamoscas de Cachemira) y Zoothera wardii (zorzal de Ward) , ambos del Himalaya se mueven tan lejos al sur como las tierras altas de Sri Lanka.
Irrupciones y dispersión
A veces circunstancias como un buena estación de cría seguida de falta de recursos alimenticios en el año siguiente llevan a una irrupción, en la cual grandes números de una especie se mueven mucho más allá de su rango normal. Bombycilla garrulus (ampelis europeo), Carduelis spinus (lúgano), y Loxia curvirostra (piquituerto común) son especies que muestran esta variación impredecible en su número anualmente.
Las zonas templadas de los continentes del sur tienen extensas áreas áridas, particularmente en Australia, y el oeste del sur de África, y movimientos provocados por el clima son comunes pero no siempre predecibles. Un par de semanas de fuertes lluvias en una parte u otra del normalmente seco centro de Australia, por ejemplo, causa el crecimiento dramático de plantas e invertebrados, atrayendo aves de todas direcciones. Esto puede suceder en cualquier época del año, y, en cualquier área determinada, puede no volver a ocurrir en una década o más, dependiendo de la frecuencia de los periodos de “El Niño” y “La Niña”.
 Merops ornatus, abejaruco australiano.
Merops ornatus, abejaruco australiano.La migración de aves es un fenómeno principalmente, aunque no enteramente, del hemisferio norte. En el hemisferio sur, la migración estacional tiende a ser mucho menos evidente. Hay varias razones para esto.
Primero, las expansiones de masas de tierra u océanos, mayormente sin interrupciones, tienden a no concentrar las migraciones por vías estrechas y evidentes, y por lo tanto un observador humano las nota menos. Segundo, al menos para aves terrestres, las regiones climáticas tienden a desvanecerse una en la otra en grandes distancias en vez de estar enteramente separadas: esto significa que en vez de hacer un largo viaje sobre un hábitat no apropiado para alcanzar un destino en particular, las especies migratorias usualmente pueden trasladarse pausada y relajadamente, alimentándose a medida que avanzan. Sin abundantes estudios de anillado no se hace obvio en estos casos que las aves vistas en un área en particular a medida que las estaciones cambian son de hecho miembros diferentes de la misma especie que van pasando gradualmente siguiendo su vía hacia el norte o el sur.
De hecho muchas especies si crían en zonas templadas del sur e invernan más al norte en los trópicos. En África, Hirundo cucullata (golondrina cabecirrufa o franjeada grande), y en Australia, Myiagra cyanoleuca (papamoscas satinado), Eurystomus orientalis (carraca verde dólar) y Merops ornatus (abejaruco arcoiris) por ejemplo, invernan muy al norte de su región de cría.
Fisiología y control
El control de la migración, su determinación cronológica y la respuesta a ésta son controlados genéticamente y parecen ser rasgos primitivos que están presentes incluso en muchas especies no migratorias. La habilidad para navegar y orientarse independientemente durante las migraciones es un fenómeno mucho más complejo que puede incluir tanto programas endógenos como aprendizaje.[20]
Bases fisiológicas
Introducción:
-La base fisiológica de la migración se basa en procesos endogenos, provocados por estímulos externos, recibidos por el SNC. (Gwinner 1986;Ketterson and Nolan1990;Healy et al.1996;Birgman1998) Como ``mensajeros´´ del proceso se encuentran las hormonas neuroendocrinas y endocrinas secretadas via hipotálamo-hipófisis. El impulso migratorio posee una fuerte componente genética, existen experimentos con lavanderas boyeras(Motacilla alba) en los que diferentes poblaciones en areas geográficas similares poseen caracteres migradores muy diferentes(Curry-Lindahl, K.1958). El proceso migratorio produce cambios importantes en la fisiologia del animal, destacando la hiperfagia, el aumento de hematocrito en sangre y algunos cambios comportamentales como el gregarismo.
Cambios que se producen en el ave:
En la fase premigratoria el ave principalmente aumenta su nivel de grasa(Blem 1990) Las grasas son su principal combustible en este proceso, se acumulan principalmente en el tejido adiposo, en los musculos y en órganos internos(George and Berger 1966). Entre las zonas más importantes de acumulo de grasa se encuentran: La clavícula, el coracoides, los costados, el abdomen, la pelvis y la zona del trasero(King and Farner 1965) Los ácidos grasos son diferentes durante la migración(predominan los ácidos grasos insaturados) que durante la fase de nidificación(predominan los ácidos grasos saturados)(Conway et al.1994). Como hemos comentado anteriormente, se acumula grasa en los músculos, pero nunca, en el corazón. La acumulación de grasas en la fase premigratoria es bien conocida desde hace muchos años por los gourmets quienes buscan a los migradores en esta época pues su carne es más fina y más rica en grasa. En función de la distancia a recorrer durante la migración el ave acumulará más o menos reservas. Meter dibujo y tabla de acumulación de grasas.
Las grasas además de aportar energia para los músculos, ayudan a la termorregulación del ave durante el proceso, además de grasas, durante la migración el ave aumenta el consumo de proteínas y carbohidratos. En la fase post-migratoria el ave experimentará un proceso hiperfágico, se ha demostrado también que durante esa fase, el ave, posee más capacidad de recuperación de reservas.
Bases neuronales y hormonas implicadas en los procesos migratorios:
No hay una glándula que intervenga por sí sola en la determinación del impulso migratorio, sino, por el contrario, todo un conjunto de glándulas endocrinas.
La hipófisis ocupa un lugar preeminente, en razón de su papel de verdadero puesto de mando del organismo, y también debido a su sensibilidad a los factores de iluminación. Además de la hipófisis se ha destacado la importancia de la tiroides(maneja la movilización de grasas en la termorregulación) y las gónadas( Rowan, W.1939, deducía de sus experimentos que para la migración era condición necesaria un desarrollo intermedio gonadal).
-Los factores ambientales condicionan el acto migratorio, afectando directamente a las glándulas anteriormente citadas, por ejemplo:
-En el caso de la tiroides, hay numerosos casos de aves que migran grandes distancias ``empujadas´´ por potentes olas de frío.
-La hipófisis se ve claramente influenciada por el fotoperiodo, cada especie, cría y migra dentro de sus margenes aptos de fotoperiodo.Existen experimentos con aves en cautividad en los cuales se pudo comprobar que tan solo con el estímulo del fotoperiodo las aves mostraban agitación orientada a sus lugares de migración.
La prolactina, la somatotropina, la hormona pancreática, la hormona hipofisial, catecolaminas e insulina tienen un papel fundamental en la acumulación de grasas, en la hipertrofia muscular y en el aumento del hematocrito(Ramenofsky and Boswell 1994).
-Catecolaminas, hormonas del crecimiento y corticosterona participan en la movilización de grasas(Ramenofsky 1990).
-La corticosterona y la testosterona es importante en la migración nocturna de aves.(Gwinner 1975).
-La melatonina tiene 1 papel fundamental en la organización de la migración y la orientación (Beldhuis et al. 1988; Schnneider et al. 1994).
Factor cronológico desencadenante
El estímulo fisiológico primario para la migración es el cambio en la longitud del día. Estos cambios están también relacionados con cambios hormonales en las aves.
En el periodo antes de la migración, muchas aves despliegan una mayor actividad o “Zugunruhe” (en alemán: inquietud migratoria) así como cambios fisiológicos como incremento de la deposición de grasas. La ocurrencia de este periodo incluso en aves enjauladas sin estímulos ambientales (por ejemplo, acortamiento del día y o disminución de la temperatura) ha indicado hacia el rol de programas endógenos con ritmos anuales en el control de la migración de las aves. Aves enjauladas muestran una dirección preferencial de vuelo que se corresponde con la dirección de la migración que ellas habrían tomado en la naturaleza, incluso cambiando sus direcciones preferenciales aproximadamente al mismo tiempo que sus coespecíficos silvestres cambian de curso.
En las especies donde existen poliginia y dimorfismo sexual acentuado, hay una tendencia a que los machos vuelvan más pronto que las hembras a los sitios de reproducción. Esto se denomina protoandria.[21] [22]
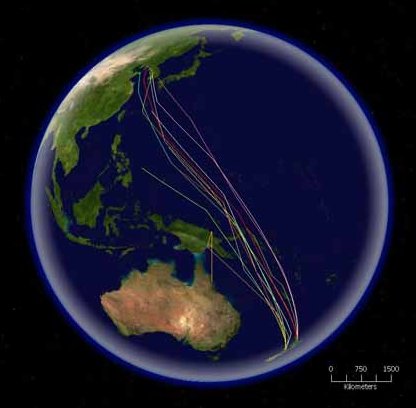
 Las rutas de especímenes de Limosa lapponica (becasina o aguja colipinta) marcados por satélite en su migración al Norte desde Nueva Zelanda. Esta especie tiene la migración sin paradas más larga que se haya registrado, alcanzando los 10.200 km.
Las rutas de especímenes de Limosa lapponica (becasina o aguja colipinta) marcados por satélite en su migración al Norte desde Nueva Zelanda. Esta especie tiene la migración sin paradas más larga que se haya registrado, alcanzando los 10.200 km.La orientación de las aves se basa en diversos sensores. En muchas especies se ha demostrado el uso un compás solar. El uso del sol para dirigirse involucra el hacer compensaciones en el cambio de su posición basadas en el tiempo. La navegación también se ha demostrado que se basa en la combinación de otras habilidades que incluyen la de detectar los campos magnéticos, el uso de puntos de referencia visuales así como pistas olfatorias.[23]
Se cree que las migrantes de largas distancias se dispersan cuando son jóvenes y se apegan a sitios de cría potenciales y a sitios de invernación favoritos. Una vez que el apego al sitio se ha producido muestran una alta fidelidad al lugar, visitándolo año tras año.[24]
La habilidad de las aves para navegar durante las migraciones no puede ser explicada completamente por la programación endógena, incluso con la ayuda de las respuestas a estímulos ambientales. La habilidad de realizar exitosamente migraciones a largas distancias probablemente sólo puede ser completamente explicada teniendo en cuenta la habilidad cognitiva de las aves para reconocer hábitats y para formar mapas mentales. El seguimiento satelital de rapaces de migración diurna como Pandion haliaetus (águila pescadora) y Pernis apivorus (halcón abejero) ha mostrado que los individuos más viejos son mejores al hacer correcciones en caso de deriva provocada por el viento.[25]
Como los patrones con ritmos anuales indican, existe un fuerte componente genético para la migración en términos de determinación cronológica y de ruta, pero éste puede ser modificado por influencias ambientales. Un ejemplo interesante donde un cambio de la ruta migratoria ha ocurrido por causas de barreras geográficas es la tendencia de algunas Sylvia atricapilla (curruca capirotada) de Europa central de migrar al oeste e invernar en Gran Bretaña en lugar de cruzar los Alpes.
Las aves migratorias pueden usar dos instrumentos electromagnéticos para hallar su punto de destino: uno que es totalmente innato y otro que depende de la experiencia. Un ave joven en su primer vuelo migratorio toma la dirección correcta de acuerdo con el campo geomagnético pero no conoce cuan lejos debe volar. Lo hace a través de un "mecanismo radical dual" dependiente de la luz y el magnetismo mediante el cual reacciones químicas, en especial de fotopigmentos sensibles a luz de longitud de onda larga, son afectadas por el campo magnético. Note que aunque esto sólo funciona durante las horas del día, no usa la posición del sol de ningún modo. En este punto el ave es similar a un niño explorador con brújula pero sin mapa, hasta que va acostumbrándose a la travesía y puede poner sus otras habilidades en uso. Con la experiencia ellas aprenden varios puntos de referencia y este "mapeo" es hecho mediante receptores basados en magnetita en el sistema trigeminal, el que le dice al ave cuan fuerte es el campo magnético. Debido a que las aves migran entre regiones norteñas y sureñas, la fortaleza del campo magnético en diferentes latitudes le permite interpretar el "mecanismo radical dual" más acertadamente, y conocer si ha llegado a su destino.[26] Investigaciones más recientes han hallado una conexión neuronal entre el ojo y el "agrupamiento N", la parte del cerebro anterior que está activa durante la orientación migratoria, sugiriendo que las aves pueden realmente ser capaces de "ver" el campo magnético de la Tierra.[27] [28]
Vagabundeo
Las aves en su migración pueden perder el rumbo y aparecer fuera de su área de distribución normal. Esto puede deberse a que se sobrepasan de su lugar de destino, por ejemplo porque vuelen más al norte que el área de cría habitual. Un mecanismo que puede originar grandes rarezas, al ocurrir que aves jóvenes vuelven atrás como vagabundos por cientos de kilómetros fuera de su rango se denomina migración revertida, en ellas falla la ejecución apropiada del programa genético.
Algunas áreas se han vuelto famosas como puntos de observación de aves debido a su localización. Por ejemplo el Parque Nacional Point Pelee en Canadá, y Cabo Spurn en Inglaterra. La deriva migratoria de aves sacadas de curso por el viento puede resultar en la "arribazón" de gran número de migrantes en sitios costeros.
Acondicionamiento del instinto migratorio
Ha sido posible enseñar una ruta de migración a una bandada de aves, por ejemplo en esquemas de reintroducción. Después de una prueba con Branta canadensis (ganso de Canadá), se usaron aviones superligeros en Estados Unidos para enseñar rutas migratorias seguras a especímenes reintroducidos de Grus americana (grulla blanca americana).[29] [30]
Factores evolutivos y ecológicos
El que una especie migre depende de un número de factores. El clima del área de cría es importante, y pocas especies pueden lidiar con los duros inviernos del interior de Canadá o el norte de Eurasia. Así es parcialmente migratorio Turdus merula (mirlo euroasiático), es migratorio en Escandinavia, pero no en el clima más templado del sur de Europa. La naturaleza del alimento principal también es importante. La mayoría de los que se especializan en comer insectos fuera de los trópicos son migrantes a largas distancias, y tienen pocas elecciones más que dirigirse al sur para pasar el invierno.
A veces los factores están finamente balanceados. Las tarabillas Saxicola rubetra (la norteña) de Europa y Saxicola maura (la siberiana) de Asia son migrantes de larga distancia invernantes en el trópico, mientras su pariente cercano Saxicola rubicola (la europea o común) es un ave residente en la mayor parte de su rango, y se mueve sólo cortas distancias desde el norte y el este más fríos. Un posible factor aquí es que las especies residentes a menudo pueden tener una nidada extra.
Estudios recientes sugieren que los paseriformes que migran largas distancias son de origen evolutivo sudamericano y africano en vez de originarse en el hemisferio norte. Son en realidad especies sureñas que van al norte a reproducirse en vez de especies norteñas que van al sur a invernar.
Análisis teóricos muestran que los desvíos y contorneos en sus rutas de vuelo que incrementan la distancia de vuelo hasta el 20% serán a menudo adaptativas desde el punto de vista aerodinámico – un ave que se llena con alimento para cruzar una amplia barrera vuela menos eficientemente. Sin embargo algunas especies muestran circuitos de rutas migratorias que reflejan una expansión histórica del rango de distribución y están lejos de ser óptimas en términos ecológicos.[31] Un ejemplo es la migración de la población continental de Catharus ustulatus (zorzalito de Swainson), que vuela muy al este atravesando Norteamérica antes de virar al sur vía la Florida para alcanzar el norte de Sudamérica; se cree que esta ruta sea una consecuencia de una expansión del rango que ocurrió hace unos 10.000 años. Los contorneos pueden ser también causados por condiciones de viento diferentes, riesgo de predación, y otros factores.
Cambio climático
Los cambios climáticos en gran escala se espera que tengan un efecto sobre la cronodeterminación de la migración, y los estudios han mostrado una variedad de efectos incluidos los cambios cronológicos en la migración, en la estación de cría.[32] así como en las declinaciones de poblaciones[33] [34]
Efectos ecológicos
La migración de las aves también ayuda al traslado de otras especies incluyendo aquellas de ectoparásitos como garrapatas y piojos,[35] los que a su vez pueden llevar microorganismos incluyendo agentes causales de enfermedades humanas. Se ha tomado interés considerable debido a la expansión global de gripe aviar, sin embargo las aves migratorias no son consideradas una amenaza mayor[36] Algunos virus que se mantienen en las aves sin efecto letal, como el West Nile virus pueden sin embargo dispersarse por la migración de las aves.[37] Las aves pueden tener también un rol en la dispersión de propágulos de plantas y plancton.[38] [39]
Algunos depredadores toman ventaja de la concentración de aves durante la migración. El murciélago Nyctalus lasiopterus (noctulo mayor) se alimenta de pájaros migratorios nocturnos.[3] Algunas aves rapaces se especializan en Charadriiformes migratorias.[40]
Técnicas de estudio
La migración de las aves ha sido estudiada por una variedad de técnicas de las cuales el anillado es la más vieja. El marcado con color, el uso de radar, el seguimiento satelital y análisis de isótopos estables de Hidrógeno (o Estroncio) son algunas de las otras técnicas usadas para estudiar las migraciones.[41]
Un método para determinar la intensidad migratoria hace uso de micrófonos apuntando hacia arriba para registrar las llamadas de contacto nocturnas de las bandadas que sobrevuelan. Éstas son luego analizadas en un laboratorio para medir tiempo, frecuencia y especies.[42]
Una técnica más antigua para cuantificar la migración involucra la observación de la cara de la luna llena y contar las siluetas de bandadas de aves a medida que vuelan en la noche.[43] [44]
Los estudios del comportamiento de orientación han sido levados a cabo tradicionalmente usando variantes de un dispositivo conocido como embudo de Emlen el cual consiste de una jaula circular cubierta arriba por vidrio o una pantalla de alambre de forma que arriba sea visible el cielo o la cúpula de un planetario o con otros estímulos ambientales controlables. El comportamiento de orientación de aves dentro de este dispositivo es estudiado cuantitativamente usando la distribución de las marcas que el ave deja en las paredes de la jaula.[45] Otros métodos usados en estudios de regreso a casa de palomas hacen uso de la dirección en que el ave desaparece en el horizonte.
Amenazas y conservación
Las actividades humanas han amenazado a muchas especies de aves migratorias. Las distancias involucradas en sus migraciones significan que ellas a menudo cruzan las fronteras de países y las medidas de conservación requieren la cooperación internacional. Varios tratados internacionales se han firmado para proteger especies migratorias incluyendo la Migratory Bird Treaty Act de 1918 de los Estados Unidos (convenio con Canadá, México, Japón y Rusia)[46] y el Acuerdo Africano-Euroasiático de Aves Acuáticas Migratorias.[47]
La concentración de aves durante la migración puede poner las especies en riesgo. Algunas de las especies migrantes más espectaculares ya se han extinguido, la más notable es Ectopistes migratorius (paloma viajera). Durante sus migraciones las bandadas eran de 1,6 km de ancho y 500 km de largo, tomando varios días para pasar y conteniendo hasta mil millones de aves.[48]
Otras áreas significativas incluyen las áreas de parada transitoria entre los territorios de cría y los de invernada.[49] Un estudio de captura y recaptura de paseriformes migrantes que tienen alta fidelidad a sus territorios de cría e invernada no mostró una asociación estricta similar con las áreas de parada transitoria.[50]
La caza a lo largo de las rutas migratorias puede causar una grave mortalidad. Las poblaciones de Grus leucogeranus (grulla siberiana) que invernan en la India declinaron debido a la caza en las rutas de transito, particularmente en Afganistán y Asia Central. Por última vez se vieron estas aves en 2002 en su sitio favorito de invernada en el Parque Nacional Keoladeo.[51]
Estructuras como las líneas de transmisión eléctrica, molinos de viento y plataformas petrolíferas de alta mar se sabe que han afectado la migración de las aves.[52] La destrucción del hábitat por el cambio en el uso de la tierra es sin embargo la amenaza más grande y los humedales de tierras bajas, que son los sitios de parada transitoria y de invernada para aves migratorias, están amenazados particularmente por el drenaje y las reclamaciones para uso humano.
Véase también
Referencias
- ↑ Peter Berthold, Hans-Günther Bauer, Valerie Westhead (2001). Bird Migration: A General Survey. ISBN 0-19-850787-9.
- ↑ Dondini, G., Vergari, S. 2000 Carnivory in the greater noctule bat (Nyctalus lasiopterus) in Italy. Journal of Zoology 251: 233-236.
- ↑ a b Popa-Lisseanu, A. G., Delgado-Huertas, A., Forero, M. G., Rodriguez, A., Arlettaz, R. & Ibanez, C. 2007. Bats' conquest of a formidable foraging niche: the myriads of nocturnally migrating songbirds. PLoS ONE 2(2): e205. URL doi:10.1371/journal.pone.0000205
- ↑ Ibáñez, C., Juste, J., García-Mudarra, J. L., Agirre-Mendi, P. T. 2001. Bat predation on nocturnally migrating birds. PNAS 98:9700-9702. Artículo completo
- ↑ a b c d Newton, I. 2008. The Migration Ecology of Birds. Elselvier. ISBN 978-0-12-517367-4
- ↑ Chan K (2001) "Partial migration in Australian landbirds: a review" Emu 101(4): 281-292)
- ↑ Boland, J. M. 1990. Leapfrog migration in North American shorebirds: intra- and interspecific examples. The Condor 92:284-290
- ↑ Hummel, D. & Beukenberg, M. (1989). Aerodynamische Interferenzeffekte beim Formationsfl ug von Vogeln. J. Ornithol. 130: 15–24.
- ↑ Cutts, C. J. & J R Speakman (1994). "Energy savings in formation flight of Pink-footed Geese". (PDF). J. Exp. Biol. 189: 251–261.
- ↑ Geroudet, P. 1954. Des oiseaux migrateurs trouves sur la glacier de Khumbu dans l'Himalaya. Nos Oiseaux 22: 254.
- ↑ Swan, L. W. 1970. Goose of the Himalayas. Nat. Hist. 79(10): 68-75.
- ↑ Dorst, J. 1963. The migration of birds. Houghton Mifflin Co., Boston. 476 p.
- ↑ Eastwood, E. and G. C. Rider. 1965. Some radar measurements of the altitude of bird flight. Br. Birds 58:
- ↑ Williams, G. G. 1950. Weather and spring migration. Auk 67: 52-65
- ↑ a b Lincoln, F. C. 1979 Migration of Birds. Fish and Wildlife Service. Circular 16.
- ↑ BTO News 258: 3, 2005
- ↑ USGS
- ↑ Schmaljohann, Heiko, Felix Liechti and Bruno Bruderer. (2007) Songbird migration across the Sahara: the non-stop hypothesis rejected! Proc Biol Sci. 274(1610):735-739 "online first" DOI: 10.1098/rspb.2006.0011
- ↑ Rattenborg, N.C., Mandt, B.H., Obermeyer, W.H., Winsauer, P.J., Huber, R.(2004) Migratory Sleeplessness in the White-Crowned Sparrow (Zonotrichia leucophrys gambelii). PLoS Biol 2(7): e212
- ↑ Helm B, Gwinner E (2006) Migratory Restlessness in an Equatorial Nonmigratory Bird. PLoS Biol 4(4): e110 doi:10.1371/journal.pbio.0040110
- ↑ Diego Rubolini, Fernando Spina and Nicola Saino (2004). "Protandry and sexual dimorphism in trans-Saharan migratory birds".. Behavioral Ecology 15(4): 592–601.
- ↑ Edwards, Darryl B.; Forbes, Mark R. (2007). "Absence of protandry in the spring migration of a population of Song Sparrows Melospiza melodia.". Ibis 149(4): 715–720.
- ↑ Walraff, H. G. 2005. Avian Navigation: Pigeon Homing as a Paradigm. Springer.
- ↑ Ketterson, E.D. and V. Nolan Jr. 1990. Site attachment and site fidelity in migratory birds: experimental evidence from the field and analogies from neurobiology. En: Bird Migration, E. Gwinner (ed.). Springer Verlag, p. 117-129 PDF
- ↑ Thorup, Kasper, Thomas Alerstam, Mikael Hake and Nils Kjelle (2003)Compensation for wind drift in migrating raptors is age dependent. Proc. R. Soc. Lond. B (Suppl.) 270, S8–S11 DOI 10.1098/rsbl.2003.0014
- ↑ Wiltschko, W., U. Munro, H. Ford & R. Wiltschko. (2006) "Bird navigation: what type of information does the magnetite-based receiver provide?" Proc. R. Soc. B. 273: 2815-20.
- ↑ Heyers D, Manns M, Luksch H, Güntürkün O, Mouritsen H (2007) A Visual Pathway Links Brain Structures Active during Magnetic Compass Orientation in Migratory Birds. PLoS ONE 2(9): e937 doi:10.1371/journal.pone.0000937 PDF
- ↑ Deutschlander, ME, Phillips, JB, Borland, SC 1999. The case for light-dependent magnetic orientation in animals J.Exp. Biol. 202:891-908 PDF
- ↑ Operation migration
- ↑ Whooping Crane Management Plan (Wisconsin)
- ↑ Alerstam, T. (2001). «Detours in bird migration» (en inglés, PDF). Journal of Theoretical Biology 209: pp. 319-331. http://orn-lab.ekol.lu.se/birdmigration/documents/pdf_articles/meglu74.pdf. Consultado el 3 de enero de 2009.
- ↑ Jenni L. & Kery M. (2003) Timing of autumn bird migration under climate change: advances in long-distance migrants, delays in short-distance migrants. Proceedings of the Royal Society of London B 270: 1467.
- ↑ Both, Christiaan; Sandra Bouwhuis, C. M. Lessells, Marcel E. Visser (2006-05-04). "Climate change and population declines in a long-distance migratory bird". Nature 441 (7089): 81–83. doi:10.1038/nature04539. ISSN 0028-0836.
- ↑ Wormworth, J. & K. Mallon (2006) Bird Species and Climate Change: The Global Status Report version 1.0. WWF. online
- ↑ Smith RP Jr, Rand PW, Lacombe EH, Morris SR, Holmes DW, Caporale DA. (1996) Role of bird migration in the long-distance dispersal of Ixodes dammini, the vector of Lyme disease. J. Infect. Dis. 174(1):221-4.
- ↑ Rappole, J.H., Hubálek, Zdenek (2006) Birds and Influenza H5N1 Virus Movement to and within North America. Emerging Infectious Diseases. 12:10 Texto completo
- ↑ Rappole, J.H., Derrickson, S.R., Hubalek, Z. (2000) Migratory Birds and Spread of West Nile Virus in the Western Hemisphere. Emerging Infectious Diseases (6)4: 319-328 Texto completo
- ↑ Figuerola, O. and A. J. Green (2002) Dispersal of aquatic organisms by waterbirds: a review of past research and priorities for future studies. Freshwater Biology 47(3), 483–494. doi:10.1046/j.1365-2427.2002.00829.x
- ↑ Cruden, R. W. 1966. Birds as Agents of Long-Distance Dispersal for Disjunct Plant Groups of the Temperate Western Hemisphere. Evolution 20(4):517-532 doi:10.2307/2406587
- ↑ Ronald C. Ydenberg, Robert W. Butler, David B. Lank, Barry D. Smith and John Ireland (2004). "Western sandpipers have altered migration tactics as peregrine falcon populations have recovered". Proc. R. Soc. Lond. B 271: 1263–1269 1263. doi:10.1098/rspb.2004.2713.
- ↑ Laura Font, Geoff M. Nowell, D. Graham Pearson, Chris J. Ottley and Stephen G. Willis (2007). "Sr isotope analysis of bird feathers by TIMS: a tool to trace bird migration paths and breeding sites". J. Anal. At. Spectrom. 22: 513. doi:10.1039/b616328a.
- ↑ Farnsworth, A., Gauthreaux, S.A., and van Blaricom, D. 2004. A comparison of nocturnal call counts of migrating birds and reflectivity measurements on Doppler radar. Journal of Avian Biology 35:365-369.
- ↑ Liechti, F. (1996) Instructions to count nocturnal bird migration by watching the full moon. Schweizerische Vogelwarte, CH-6204 Sempach, Switzerland.
- ↑ Lowery, G.H. (1951) A quantitative study of the nocturnal migration of birds. University of Kansas Publications, Museum of Natural History 3, 361-472
- ↑ Emlen, S. T. and Emlen, J. T. (1966). A technique for recording migratory orientation of captive birds. Auk 83, 361–367.
- ↑ Migratory bird Treaty 16 USC 703-711; 40 Stat. 755
- ↑ African-Eurasian Migratory Waterbird Agreement
- ↑ Schauenberg, P. et al. (1979). Fichero Safari Club (Paloma viajera). Madrid: S.A.P.E.. ISBN 84-7461-167-9.
- ↑ Shimazaki, Hiroto; Masayuki Tamura and Hiroyoshi Higuchi (2004) Migration routes and important stopover sites of endangered oriental white storks (Ciconia boyciana) as revealed by satellite tracking. Mem. Natl Inst. Polar Res., Spec. Issue, 58:162–178 PDF
- ↑ Catry, P., Encarnacao, V., Araujo, A., Fearon, P., Fearon, A., Armelin, M. & Delaloye, P. 2004. Are long-distance migrant passerines faithful to their stopover sites? Journal of Avian Biology, 35, 170–181.
- ↑ Siberian Crane fact sheet
- ↑ Fish and Wildlife Service- Bird Mortality Fact sheet
Bibliografía
- Berthold, Peter (2001) (en inglés). Bird Migration: A General Survey. (Segunda edición). Oxford University Press. ISBN 0-19-850787-9.
- Dingle, Hugh (1996) (en inglés). Migration: The Biology of Life on The Move.. Oxford University Press.
- Weidensaul, Scott (1999.) (en inglés). Living On the Wind: Across the Hemisphere With Migratory Birds.. Douglas & McIntyre.
- Edición dedicada de Philosophical Transactions B sobre la Adaptación al Ciclo Anual. Algunos artículos tienen acceso libre
- Curry-Lindahl, K.(1958)Internal timer and spring migration in an ecuatorial migrante, the Yellow Wagtail(Motacilla flava)Arkiv Zool., 11(33):541-557.
- Blem, C.R. (1990)Avian energy storage. In Current ornithology(ed. M. Power), pp.59-113-Plenum Press, New York.
- George, J. C. y Berger, a.J.(1966)Avian myology . Academic Press. Nueva York, Londres.
- King, J.R. and Farner, D.S.(1965). Studies of fat deposition in migratory birds.Ann.New York Acad. Sci. 131, 422-40.
- Conway et al.(1994).Seasonal changes in fatty acid composition of the wood trush. Cóndor 96,791-4.
- Rowan, W.(1939).Light and seasonal reproduction in animals.Biol.Rev.,13:374-402.
- Ramenofsky, M. and Boswell, T. (1994).Regulation of feeding cycles during migration: a possible role for insulin. Abstracts of 21st International Ornithological Congress.J.Ornithol.135,Sonderh.,67.
- Ramenofsky, M. (1990)Fat storage and fat metabolism in relation to migration. In Bird migration : the physiology and ecophysiology (ed. E. Gwinner), pp. 214-31. Springer – Verlag , Berlin.
- Gwinner (1975).Effects of season and external testosterone on the freerunning circadian activity rhythm of European starlings(Sturnus vulgaris).J.Comp.Physiol.103,315-28.
- Beldhuis, H.J.A. et al. (1988)Melatonin and the circadian rhythms of feeding and perch-hopping in the European Starling, Sturnus vulgaris. J.Comp.Physiol.A 164, 7-14.
- Schnneider, T. et al. (1994)Melatonin is crucial for the migratory orientation of Pied flycatchers (Ficedula hipoleuca Pallas).J.Exper. Biol.194,255-62.
- Gwinner (1986).Circannual rhythms. Springer- Verlag, Berlin.
- Ketterson and Nolan(1990).Site attachment and site fidelity: experimental evidence from the fiel dand analogies from neurobiology . In Bird migration: the physiology and ecophysiology (ed. E.Gwinner), pp. 117-29.Springer- Verlag, Berlin.
- Healy et al.(1996) Hippocampal volume in migratory and non-migratory warblers : effects of age and experience . Behav. Brain Res. 81, 61-8.
- Bingman, V.P.(1998).Spatial representations and homing pigeon navigation. In Spatial representation in animals(ed. S. Healy), pp.69-85.Oxford University Press.
- Bird Migration, a general survey, second edition. Peter Berthold.
- Ornithology. Frank b. Gill.
- Las aves en su medio ambieente. Dorst
- The migration ecology of birds. Ian Newton.
Enlaces externos
- Migration Ecology Group, Lund University, Sweden
- MIGRATE: Migration Interest Group: Research Applied Toward Education, USA – Grupo de interés sobre migración: investigación aplicada hacia la educación, Estados Unidos
- Trektellen.nl – Conteos de migración y registros de anillamiento de Holanda, Bélgica, Gran Bretaña, Francia, Alemania, España y Portugal
- Bird Research by Science Daily- incluye varios artículos sobre la migración de las aves
- The Nature Conservancy's Migratory Bird Program- El programa de aves migratorias del Nature Conservancy
- The Compasses of Birds – una revisión de la Science Creative Quarterly
- Soaring with Fidel – sigue la migración anual de las águilas pescadoras (Pandion haliaetus) desde Cabo Cod hasta Cuba y Venezuela
- Birder's Journal: A Morning With Migrants Nationalgeographic.com
- Bat predation on migrating birds – Predación de murciélagos sobre aves migratorias.
- Conteos de migración en el área del Estrecho de Gibraltar y España.












Wikimedia foundation. 2010.