- Tyrannosauroidea
-
 Tyrannosauroidea
TyrannosauroideaRango temporal: Jurásico superior – Cretácico Superior 
Appalachiosaurus.Clasificación científica Reino: Animalia Filo: Chordata Clase: Sauropsida Superorden: Dinosauria Orden: Saurischia Suborden: Theropoda (sin clasif.): Coelurosauria Superfamilia: †Tyrannosauroidea
Osborn, 1906Géneros Ver el texto.
Los tiranosauroideos (Tyrannosauroidea, nombre que significa "[forma de] lagartos tiranos") es una superfamilia (o clado) de dinosaurios terópodos celurosaurios [que incluye a la familia Tyrannosauridae así como a varios parientes más basales. Los tiranosauroideos vivieron en el supercontinente de Laurasia comenzando en el período Jurásico. A finales del período Cretácico, los tiranosauroideos eran los grandes depredadores en el Hemisferio Norte, culminando en el gigantesco Tyrannosaurus, que da nombre al grupo. Los fósiles de tiranosauroideos se han recuperado en lo que ahora son las masas continentales de Norteamérica, Europa, Asia y posiblemente Australia.
Los tiranosauroideos eran carnívoros bípedos, como muchos terópodos, y se caracterizaban por numerosos rasgos esqueléticos, especialmente del cráneo y la pelvis. A principio de su existencia, los tiranosauroideos eran pequeños depredadores con largos brazos de tres dedos. Sus descendientes en el Cretácico Superior se volvieron mucho más grandes, incluyendo a algunos de los mayores depredadores terrestres que hayan existido, pero muchos de estos géneros tenían brazos proporcionalmente pequeños de sólo dos dedos. Plumas primitivas se han hallado en Dilong, un tiranosauroideo temprano de China, y pudo haber estado presente en otros tiranosauroideos también. Prominentes crestas y protuberancias óseas de diferentes tamaños y formas en los cráneos de muchos tiranosauroideos pueden haber servido para funciones de exhibición.
Contenido
Descripción
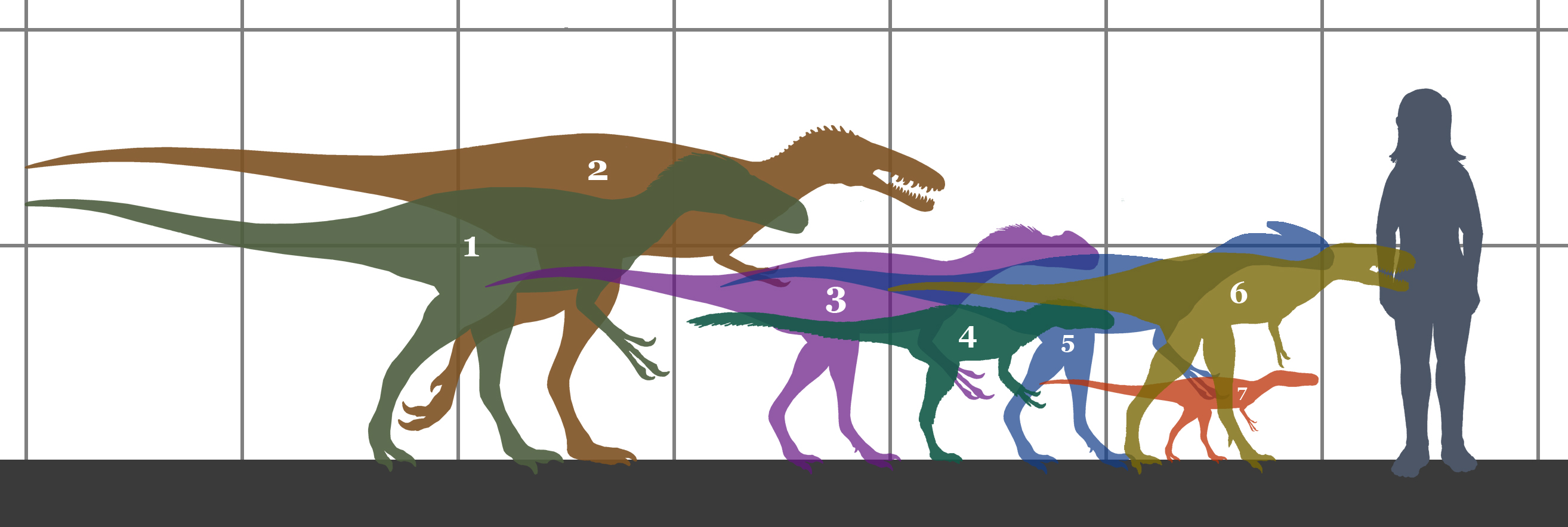
 Tamaño de algunos de los géneros más pequeños del grupo, comparados a un humano.
Tamaño de algunos de los géneros más pequeños del grupo, comparados a un humano.
Los tiranosauroides eran muy variados en tamaño, aunque tuvieron una cierta tendencia al aumento de tamaño con el tiempo. Los primeros tiranosauroides eran animales pequeños.[1] Un espécimen de Dilong, casi totalmente crecido, medía 1.6 metros de longitud,[2] y un Guanlong adulto medía 3 metros de largo.[3] Dientes de rocas del Cretácico Inferior (140 to 136 millones de años) de Hyogo, Japón, parecen provenir de un animal de aproximadamente 5 metros de largo, un posible indicio de un temprano incremento de tamaño en el linaje.[4] Un Eotyrannus inmaduro medía cerca de 4 metros de largo,[5] y un Appalachiosaurus subadulto se ha estimado en más de 6 metros de largo,[1] indicando que ambos géneros alcanzaban tamaños mayores. Los tiranosáuridos del Cretácico Tardío oscilaban desde los 8-9 metros de Albertosaurus y Gorgosaurus a los más de 12 metros de longitud de Tyrannosaurus, que pudo haber pesado más de 6.400 kilogramos.[1] Una revisión de 2010 del estado del arte concluyó que los tiranosaurios fueron "de tamaño pequeño a medio" durante sus primeros 80 millones de años mientras que fueron "algunos de los mayores carnívoros terrestres que hayan vivido" en sus últimos 20 millones de años de existencia.[6] [7]

Los cráneos de los primeros tiranosauroides eran largos, bajos y de constitución ligera, similares a los de otros celurosaurios, mientras que las formas tardías tenían cráneos más altos y macizos. En el primitivo linaje de los proceratosáuridos, además solían tener crestas decorativas en el morro, que eran muy llamativas en el Guanlong de China. A pesar de las diferencias en forma, se hallan ciertas características craneales en todos los tiranosauroides conocidos. El hueso premaxilar es muy alto, despuntando el frente del hocico, un rasgo que evolucionaron convergentemente con los abelisáuridos. Los huesos nasales están fusionados juntos, arqueados levemente hacia arriba y frecuentemente adornados con rugosidades en su superficie superior. Los dientes premaxilares en el frente de la mandíbula superior poseen una forma diferente a la del restos de los dientes, siendo mucho menores y con una forma de "D" en vista transversal. En la mandíbula inferior, una cresta prominente en el hueso suprangular se extiende de costado desde justo debajo de la articulación de la mandíbula, excepto en el primitivo Guanlong.[1] [2] [3]
Los tiranosauroides tenían un cuello en forma de S y largas colas, como muchos otros terópodos. Los géneros primitivos tenían largos miembros delanteros, siendo cerca del 60% de la longitud del miembro posterior en Guanlong, con los típicos tres dígitos de los celurosaurios.[3] Los largos brazos persistieron al menos a través del Cretácico Inferior, como es el caso de Eotyrannus,[5] pero se desconoce en Appalachiosaurus.[8] Los más derivados tiranosáuridos tienen brazos muy reducidos en tamaño, siendo el más extremo ejemplo Tarbosaurus de Mongolia, en el cual su húmero sólo era un cuarto de la longitud del fémur.[1] El tercer dedo del miembro delantero también se redujo con el paso del tiempo. Este dígito no estaba reducido en el basal Guanlong,[3] pero en Dilong era significativamente más delgado que los otros dos dedos.[2] Eotyrannus aún tenía tres dedos funcionales en sus manos,[5] pero los tiranosáuridos sólo tenían dos, si bien remanentes vestigiales del tercero son hallados en algunos especímenes.[9] Como en muchos celurosaurios, el segundo dígito era el más grande, incluso cuando el tercero no estaba presente.
Rasgos característicos de la pelvis de los tiranosauroides incluyen una muesca cóncava en el extremo superior frontal del iliaco, una definida cresta vertical afilada en la superficie externa del iliaco, que se extiende hacia arriba desde el acetábulo (fosa de la cadera), y una gran "bota" pélvica en el extremo del pubis, más de la mitad de larga que el propio eje del pubis.[1] Estas características son halladas en todos los tiranosauroides conocidos, incluyendo a miembros basales como Guanlong[3] y Dilong.[2] El pubis no es conocido en Aviatyrannis o Stokesosaurus pero ambos muestran características típicas de los tiranosauroides en el iliaco.[10] Los miembros posteriores de todos los tiranosauroides, como muchos terópodos, tenían cuatro dedos, aunque el primer dedo (el hallux) no estaba en contacto con el suelo. Las patas de los tiranosauroides son relativamente largos con respecto al tamaño del cuerpo comparados con la mayoría de los terópodos, y muestran proporciones propias de animales corredores, incluyendo tibias y metatarsos alargados.[1] Estas proporciones persisten incluso en los mayores adultos de Tyrannosaurus,[11] a pesar de su posible incapacidad de correr a gran velocidad.[12] El tercer metatarso de los tiranosáuridos estaba comprimido en la parte superior entre el segundo y el cuarto, formando una estructura conocida como el arctometatarso.[1] El arctometatarso estaba también presente en Appalachiosaurus[8] pero no es claro si fue hallado en Eotyrannus[5] o Dryptosaurus.[13] Esta estructura era compartida por los ornitomímidos, troodóntidos y cenagnátidos,[14] pero no en los tiranosauroides basales como Dilong, indicando una evolución convergente.[2]
Sistemática
Tyrannosaurus fue nombrado por Henry Fairfield Osborn en 1905, junto con la familia Tyrannosauridae.[15] El nombre se deriva de los términos del griego antiguo τυραννος/tyrannos ('tirano') y σαυρος/sauros ('lagarto' o 'reptil'). El nombre de la superfamilia Tyrannosauroidea fue publicado en un artículo de 1964 por el paleontólogo británico Alick Walker.[16] El sufijo -oidea, comúnmente usado en los nombres de superfamilias de animales, se deriva del griego ειδος/eidos ('forma').[17]
Los científicos han comúnmente entendido a Tyrannosauroidea como el grupo que incluye a los tiranosáuridos y a sus ancestros immediatos.[16] [18] Sin embargo, con la llegada de la taxonomía filogenética en la paleontología de vertebrados, el clado ha recibido varias definiciones más explícitas. La primera fue hecha por Paul Sereno en 1998, en la que Tyrannosauroidea fue definido como un taxón basado en ramas que incluye todas las especies que comparten el más reciente ancestro común con Tyrannosaurus rex que con las aves Neornithes.[19] Para hacer a la familia más exclusiva, Thomas Holtz la redefinió en 2004 para incluir a todas las especies más cercanamente relacionadas a Tyrannosaurus rex que a Ornithomimus velox, Deinonychus antirrhopus o a Allosaurus fragilis.[1] Sereno publicó una nueva definición en 2005, usando a Ornithomimus edmontonicus, Velociraptor mongoliensis y a Troodon formosus como especificadores externos.[20] La definición de Sereno fue adoptada en una revisión de 2010.[6]
Taxonomía
 Una pintura de 1897 de Laelaps (ahora conocido como Dryptosaurus) por Charles R. Knight. Dryptosaurus fue un tiranosauroide basal relicto del este de Norteamérica.
Una pintura de 1897 de Laelaps (ahora conocido como Dryptosaurus) por Charles R. Knight. Dryptosaurus fue un tiranosauroide basal relicto del este de Norteamérica. Tyrannosaurus rex, uno de los últimos, y también el mayor de los tiranosauroides conocidos.
Tyrannosaurus rex, uno de los últimos, y también el mayor de los tiranosauroides conocidos.Clasificación
SUPERFAMILIA TYRANNOSAUROIDEA
- Alectrosaurus (Cretácico Superior, Mongolia)
- Appalachiosaurus (Cretácico Superior, este de Norteamérica)
- Aviatyrannis (Jurásico Superior, Portugal)
- ?Bagaraatan (Cretácico Inferior, Mongolia)
- Bistahieversor (Cretácico Superior, Nuevo México, Estados Unidos)
- Dilong (Cretácico Inferior, este de China)
- ?Diplotomodon[21] (Cretácico Superior, Nueva Jersey, Estados Unidos)
- Eotyrannus (Cretácico Inferior, Inglaterra)
- ?Iliosuchus (Jurásico Medio, Inglaterra)
- ?Labocania (Cretácico Superior, occidente de México)
- Stokesosaurus (Jurásico Superior, oeste de Norteamérica)
- Xiongguanlong (Cretácico Inferior, China central)
- ?Familia Coeluridae[22]
- Coelurus (Jurásico Superior, oeste de Norteamérica)
- Tanycolagreus (Jurásico Superior, oeste de Norteamérica)
- Familia Dryptosauridae[23]
- Dryptosaurus (Cretácico Superior, este de Norteamérica)
- Familia Proceratosauridae[24]
- Guanlong (Jurásico Medio, oeste de China)
- Kileskus (Jurásico Medio, Rusia central)[25]
- Proceratosaurus (Jurásico Medio, Inglaterra)
- Sinotyrannus (Cretácico Inferior, este de China)[6]
- Familia Tyrannosauridae
- Albertosaurus (Cretácico Superior, oeste de Norteamérica)
- Alioramus (Cretácico Superior, Mongolia)
- Aublysodon? (Cretácico Superior, oeste de Norteamérica)
- Daspletosaurus (Cretácico Superior, oeste de Norteamérica)
- Gorgosaurus (Cretácico Superior, oeste de Norteamérica)
- Nanotyrannus (Cretácico Superior, oeste de Norteamérica)
- Raptorex? (Cretácico Superior?, Mongolia?)
- Tarbosaurus (Cretácico Superior, Mongolia)
- Teratophoneus (Cretácico Superior, oeste de Norteamérica)
- Tyrannosaurus (Cretácico Superior, oeste de Norteamérica)
- Zhuchengtyrannus (Cretácico Superior, China)
Filogenia
Aunque los paleontólogos han reconocido siempre a la familia Tyrannosauridae, sus ancestros han sido materia de un amplio debate. A lo largo de casi todo el siglo XX, los tiranosáuridos fueron comúnmente aceptados como miembros del grupo Carnosauria, el cual incluía a casi todos los demás terópodos grandes.[26] [27] Dentro de este grupo, los alosáuridos eran frecuentemente considerados como los ancestros de los tiranosáuridos.[18] [28] Sin embargo, a principios de la década de 1990, los análisis cladísticos empezaron a situar a los tiranosáuridos dentro del grupo Coelurosauria,[14] [29] haciendo eco de hipótesis que habían sido publicadas en la década de 1920.[30] [31] Los tiranosáuridos ahora son universalmente considerados como grandes celurosaurios.[1] [3] [22] [32] [33] [34] [35]
Cladogramas de los Tyrannosauroidea basales
Holtz 2004[1] Coelurosauria void Tyrannosauroidea void void void void void
Carr et al. 2005[8] Tyrannosauroidea void void
Xu Xing et al. 2006[3] Coelurosauria void Tyrannosauroidea void void Dilong
Brusatte et al. 2010[6] Tyrannosauroidea Proceratosauridae void void void void Dilong
void void void void void void void Bistahieversor
En 1994, Holtz agrupó a los tiranosauroides con los elmisáuridos, ornitomimosaurios y los troodóntidos dentro de un clado de celurosaurios llamado Arctometatarsalia basado en una estructura común del tobillo en la que el segundo y el cuarto metatarsal se encuentran cerca de los huesos del tarso, cubriendo el tercer metatarso cuando es visto de frente.[14] Tiranosauroides basales como Dilong, sin embargo, carecen de estos tobillos arctometatarsales, indicando que esta característica evolucionó de manera independiente.[2] Arctometatarsalia ha sido desmantelado y ya no es usado por la mayoría de los paleontólogos, al tiempo que los tiranosauroides se consideran usualmente como celurosaurios basales por fuera del grupo Maniraptoriformes.[1] [33] [35] Un reciente análisis halló que la familia Coeluridae, que incluye a los géneros norteamericanos de finales del Jurásico Coelurus y Tanycolagreus, eran el taxón hermano de Tyrannosauroidea.[22]
El tiranosauroide más basal conocido de restos del esqueleto completo es Guanlong.[3] Otros taxones primitivos como Stokesosaurus y Aviatyrannis, son conocidos de material mucho menos completo.[10] El mejor conocido Dilong es considerado como algo más derivado que Guanlong y Stokesosaurus.[2] [3] Dryptosaurus, que casi siempre ha sido un género difícil de clasificar, ha sido recuperado como un tiranosauroide basal en varios análisis recientes, algo más distantemente relacionado a Tyrannosauridae que Eotyrannus y Appalachiosaurus.[1] [8] [36] Alectrosaurus, un género pobremente conocido de Mongolia, es definitivamente un tiranosauroide pero sus relaciones exactas no son claras.[1] Otros taxones han sido considerados como posibles tiranosauroides por varios autores, incluyendo a Bagaraatan, Labocania y una especie erróneamente referida a Chilantaisaurus, "C." maortuensis;[1] esta última sería posteriormente reclasificada como un género propio de carcarodontosáurido, Shaochilong.[37] Siamotyrannus del Cretácico Inferior de Tailandia fue originalmente descrito como un tiranosáurido primitivo,[38] pero hoy en día es considerado como un carnosaurio.[33] [39] Iliosuchus tiene una cresta vertical en el hueso iliaco que recuerda al de los tiranosauroides y puede ser de hecho el más antiguo miembro conocido de la superfamilia, pero no hay suficiente material para estar seguro.[10] [39]
Distribución
Los tiranosauroides vivieron en el supercontinente de Laurasia, el cual se separó de Gondwana en el Jurásico Medio, así como en los continentes del hemisferio norte, que emergieron de la división de Laurasia más adelante en el Mesozoico. Los primeros tiranosauroides vivieron a fines del Jurásico, incluyendo a Guanlong del noroeste de China,[3] Stokesosaurus del occidente de Estados Unidos y Aviatyrannis de Portugal. Algunos fósiles actualmente referidos a Stokesosaurus pueden pertenecer a Aviatyrannis, dadas las grandes similitudes entre las faunas de dinosaurios de Portugal y Norteamérica en este tiempo. Si Iliosuchus del Jurásico Medio de Inglaterra es de hecho un tiranosauroide, podría ser el más antiguo género conocido y podría sugerir que la superfamilia se originó en Europa.[10]
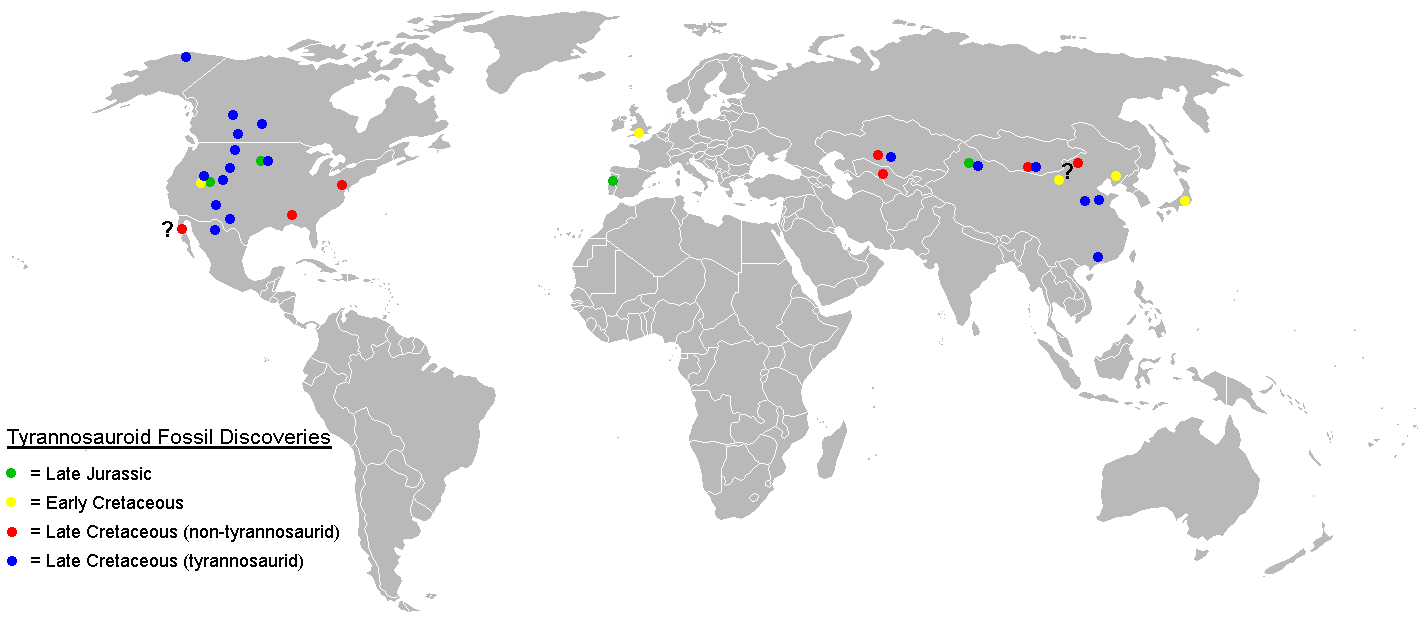
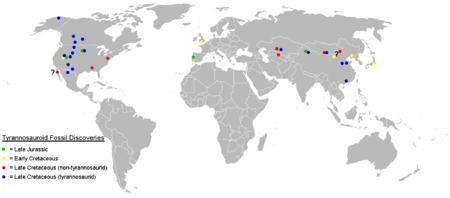 Fósiles confirmados de tiranosauroides sólo se han hallado en los continentes del norte, con posibles fósiles de tiranosauroides basales reportados de Australia. Tiranosauroides del Cretácico Tardío sólo se conocen de Norteamérica y Asia.
Fósiles confirmados de tiranosauroides sólo se han hallado en los continentes del norte, con posibles fósiles de tiranosauroides basales reportados de Australia. Tiranosauroides del Cretácico Tardío sólo se conocen de Norteamérica y Asia.Tiranosauroides del Cretácico Inferior se hallan en los tres continentes del Hemisferio Norte. Eotyrannus de Inglaterra[5] y Dilong del noreste de China[2] son los únicos dos géneros nombrados de esta época, si bien dientes premaxilares de tiranosauroide se hallan en la formación Cedar Mountain del Cretácico Inferior en Utah[40] y el Grupo Tetori de Japón.[41]
A mediados del Cretácico, ya no se encuentran fósiles de tiranosauroidea en Europa, lo que sugiere que hubo una extinción local en ese continente.[1] Dientes de tiranosauroides y posibles cuerpos fosilizados son conocidos de Norteamérica en la formación Dakota,[40] y en formaciones de Kazakstán, Tayikistán y Uzbekistán,[42] de mediados del Cretácico. Los primeros restos indiscutibles del tiranosáuridos aparecen en el Campaniano ya en el Cretácico Superior de Norteamérica y Asia. Se reconocen dos subfamilias. Los albertosaurinos son solo conocidos de América del Norte, mientras que los tiranosaurinos se encuentran en ambos continentes.[1] Fósiles de tiranosáuridos de han descubierto en Alaska, que pudo haber servido de puente terrestre permitiendo la dispersión entre ambos continentes.[43] Tiranosauroides no tiranosáuridos como Alectrosaurus y posiblemente Bagaraatan fueron contemporáneos de los tiranosáuridos en Asia, pero estaban ausentes en el oeste de Norteamérica.[1] El oriente de Norteamérica se encontraba aislado por el Mar Niobraran a mediados del Cretácico de la parte occidental del continente. La ausencia de tiranosáuridos en la parte oriental del continente sugiere que esa familia evolucionó después de la aparición de la vía marítima, permitiendo a tiranosauroides basales como Dryptosaurus y Appalachiosaurus sobrevivir en el este como una población relicta hasta el final del Cretácico.[8]
Tiranosauroides basales pueden haber estado presentes en lo que ahora es el sureste de Australia durante el Aptiano, en el Cretácico Inferior. El ejemplar NMV P186069, un pubis parcial (uno de los huesos de la cadera) con una distintiva forma similar a la de un tiranosauroide, fue descubierto en el yacimiento de Dinosaur Cove en Victoria, indicando que probablemente los tiranosauroides no estaban limitados a los continentes del norte, como anteriormente se había pensado.[44]
Paleobiología
Plumas
Largas estructuras filamentosas se han preservado con restos del esqueleto de numeroso celurosaurios del Cretácico Inferior en la formación Yixian y otras formaciones geológicas cercanas de Liaoning, China.[45] Estos filamentos se han interpretado usualmente como "protoplumas," homólogas con las plumas ramificadas propias de las aves y algunos terópodos no avianos,[46] [47] si bien se han propuesto otras hipótesis.[48] Un esqueleto de Dilong fue descrito en 2004 e incluía el primer ejemplo de "protoplumas" en un tiranosauroide. Similares al plumón de las aves modernas, las protoplumas halladas en Dilong eran ramificadas pero no penáceas, y pueden haberse usado para aislación térmica.[2]
La presencia de protoplumas en los tiranosauroideos basales no es sorprendente, dado que se sabe que eran características de los celurosaurios, y se hallan en otros géneros basales como Sinosauropteryx,[46] as well as all more derived groups.[45] Sin embargo, raras impresiones fosilizadas de piel de grandes tiranosáuridos carecen de plumas, mostrando en cambio que poseían una cubierta de escamas.[49] Es posible que las protoplumas estuvieran presentes en áreas del cuerpo que no preservaron impresiones de piel. Otra posibilidad es que hubiera una pérdida secundaria de las protoplumas en los grandes tiranosáuridos en analogía con la pérdida de pelo en los grandes mamíferos modernos como los elefantes, que tiene una escasa proporción de área superficial en relación a su volumen lo que ralentiza la transferencia de calor, haciendo que el aislamiento con una capa de pelo sea innecesario.[2]
Crestas de la cabeza
Crestas y protuberancias óseas son halladas en los cráneos de muchos terópodos, incluyendo a numerosos tiranosauroides. La más elaborada se encuentra en el Guanlong, en el cual los huesos nasales una única gran cresta que corría a lo largo de la parte media del cráneo, desde el frente hasta la parte posterior. Esta cresta estaba atravesada por varios grandes forámenes (aberturas) que reducían su peso.[3] Una cresta menos prominente es hallada en Dilong, en la que bordes bajos y paralelos corían a lo largo del cráneo de cada lado del mismo, apoyándose en los huesos nasales y lacrimales. Estos bordes se curvaban hacia adentro y se encontraban justo detrás de las narinas, dándole a la cresta una forma de letra Y.[2] Los huesos nasales fusionados de los tiranosáuridos tienen frecuentemente una textura muy rugosa. Alioramus, un posible tiranosáurido de Mongolia, llevaba una única hilera de cinco bultos óseos sobre el hueso nasal; una hilera similar de bultos más bajos está presente en el cráneo de Appalachiosaurus, así como en algunos especímenes de Daspletosaurus, Albertosaurus y Tarbosaurus.[8] En Albertosaurus, Gorgosaurus y Daspletosaurus, además poseen un prominente "cuerno" en frente de cada ojo sobre el hueso lacrimal. El cuerno lacrimal está ausente en Tarbosaurus y Tyrannosaurus, que en cambio tenían una cresta en forma de media luna detrás de cada ojo en el hueso postorbital.[1]
Estas crestas craneales pueden haberse usado para exhibirse, quizás para reconocimiento de miembros de la misma especie o para cortejarse.[1] Esto sería un ejemplo del principio de handicap, como pudo presentarse en el caso de Guanlong, en el cual su gran y delicada cresta puede haber sido un estorbo para cazar, asumiendo que seguramente era un depredador activo. Si un individuo era sano y exitoso cazando a pesar de su frágil cresta, esto podría indicar las buenas cualidades del individuo sobre otros con crestas menores. De forma similar a la inmanejable cola de un pavo real o a las desmesuradas astas de un Megaloceros, la cresta de Guanlong puede haber evolucionado a través de la selección sexual, proveyéndole una ventaja en el cortejo lo cual hubiera compensado cualquier decrecimiento en las habilidades de caza.[3]
Reproducción
Fósiles de tiranosaurios neonatos se han documentado en la literatura científica.[50]
Referencias
- ↑ a b c d e f g h i j k l m n ñ o p q r s Holtz, Thomas R. (2004). «Tyrannosauroidea». En Weishampel, David B.; Dodson, Peter; & Osmólska, Halszka (eds.).. The Dinosauria (Second edición). Berkeley: University of California Press. pp. 111–136. ISBN 0-520-24209-2.
- ↑ a b c d e f g h i j k Xu Xing, X; Norell, Mark A.; Kuang Xuewen; Wang Xiaolin; Zhao Qi; & Jia Chengkai. (2004). «Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids». Nature 431 (7009): pp. 680–684. doi:. PMID 15470426.
- ↑ a b c d e f g h i j k l Xu Xing, X; Clark, James M.; Forster, Catherine A.; Norell, Mark A.; Erickson, Gregory M.; Eberth, David A.; Jia Chengkai; & Zhao Qi. (2006). «A basal tyrannosauroid dinosaur from the Late Jurassic of China». Nature 439 (7077): pp. 715–718. doi:. PMID 16467836.
- ↑ «Tyrannosaurus ancestor's teeth found in Hyogo». The Japan Times (21-06-2009). Consultado el 22-08-2009.
- ↑ a b c d e Hutt, Stephen; Naish, Darren; Martill, David M.; Barker, Michael J.; & Newberry, Penny. (2001). «A preliminary account of a new tyrannosauroid theropod from the Wessex Formation (Early Cretaceous) of southern England». Cretaceous Research 22 (2): pp. 227–242. doi:.
- ↑ a b c d Brusatte, S.L.; Norell, M.A.; Carr, T.D.; Erickson, G.M.; Hutchinson, J.R.; Balanoff, A.M.; Bever, G.S.; Choiniere, J.N.; Makovicky, P.J.; and Xu, X. (2010). «Tyrannosaur paleobiology: new research on ancient exemplar organisms». Science 329 (5998): pp. 1481−1485. doi:. PMID 20847260. Bibcode: 2010Sci...329.1481B.
- ↑ Handwerk, B.. «Tyrannosaurs were human-size for 80 million years», 16 de septiembre de 2010. Consultado el 17 de septiembre de 2010.
- ↑ a b c d e f Carr, Thomas D.; Williamson, Thomas E.; & Schwimmer, David R. (2005). «A new genus and species of tyrannosauroid from the Late Cretaceous (middle Campanian) Demopolis Formation of Alabama». Journal of Vertebrate Paleontology 25 (1): pp. 119–143. doi:. http://www.bioone.org/perlserv/?request=get-abstract&doi=10.1671%2F0272-4634(2005)025%5B0119%3AANGASO%5D2.0.CO%3B2.
- ↑ Quinlan, Elizibeth D.; Derstler, Kraig; & Miller, Mercedes M. (2007). «Anatomy and function of digit III of the Tyrannosaurus rex manus». Geological Society of America Annual Meeting — Abstracts with Programs: pp. 77. http://gsa.confex.com/gsa/2007AM/finalprogram/abstract_132345.htm. [abstract only]
- ↑ a b c d Rauhut, Oliver W.M. (2003). «A tyrannosauroid dinosaur from the Upper Jurassic of Portugal». Palaeontology 46 (5): pp. 903–910. doi:.
- ↑ Brochu, Christopher R. (2003). «Osteology of Tyrannosaurus rex: insights from a nearly complete skeleton and high-resolution computed tomographic analysis of the skull». Society of Vertebrate Paleontology Memoirs 7: pp. 1–138. doi:. http://www.vertpaleo.org/publications/memoirs.cfm.
- ↑ Hutchinson, John R.; & Garcia, Mariano . (2002). «Tyrannosaurus was not a fast runner». Nature 415 (6875): pp. 1018–1021. doi:. PMID 11875567.
- ↑ Carpenter, Kenneth; Russell, Dale A.; Baird, Donald; & Denton, Robert (1997). «Redescription of the holotype of Dryptosaurus aquilunguis (Dinosauria: Theropoda) from the Upper Cretaceous of New Jersey». Journal of Vertebrate Paleontology 17 (3): pp. 561–573. doi:. http://www.vertpaleo.org/publications/jvp/17-561-573.cfm.
- ↑ a b c Holtz, Thomas R. (1994). «The phylogenetic position of the Tyrannosauridae: implications for theropod systematics». Journal of Palaeontology 68 (5): pp. 1100–1117. http://jpaleontol.geoscienceworld.org/cgi/content/abstract/68/5/1100.
- ↑ Osborn, Henry F. (1905). «Tyrannosaurus and other Cretaceous carnivorous dinosaurs». Bulletin of the American Museum of Natural History 21: pp. 259–265. doi:. http://digitallibrary.amnh.org/dspace/handle/2246/1464.
- ↑ a b Walker, Alick D. (1964). «Triassic reptiles from the Elgin area: Ornithosuchus and the origin of carnosaurs». Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 248: pp. 53–134. doi:. Bibcode: 1964RSPTB.248...53W.
- ↑ Liddell, Henry G.; & Scott, Robert (1980). Greek-English Lexicon (Abridged edición). Oxford: Oxford University Press,. ISBN 0-19-910207-4.
- ↑ a b Bonaparte, José F.; Novas, Fernando E.; & Coria, Rodolfo A. (1990). «Carnotaurus sastrei Bonaparte, the horned, lightly built carnosaur from the Middle Cretaceous of Patagonia». Contributions in Science (Natural History Museum of Los Angeles County 416: pp. 1–42.
- ↑ Sereno, Paul C. (1998). «A rationale for phylogenetic definitions, with application to the higher-level taxonomy of Dinosauria». Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 210 (1): pp. 41–83.
- ↑ Sereno, Paul C. (2005). «Stem Archosauria — TaxonSearch, Version 1.0». Consultado el 10-12-2007.
- ↑ Molnar, (1990). "Problematic Theropoda: "Carnosaurs"." Pp. 306-317 in Weishampel et al. (eds.), The Dinosauria. Berkeley: University of California Press.
- ↑ a b c Senter, Phil (2007). «A new look at the phylogeny of Coelurosauria (Dinosauria, Theropoda)». Journal of Systematic Palaeontology 5 (4): pp. 429–463. doi:.
- ↑ Carpenter, Ken & Russell, Dale A, Donald Baird, and R. Denton (1997). «Redescription of the holotype of Dryptosaurus aquilunguis (Dinosauria: Theropoda) from the Upper Cretaceous of New Jersey.». Journal of Vertebrate Paleontology 17 (3): pp. 561–573. doi:.
- ↑ Rauhut, Oliver W. M.; Milner, Angela C.; and Moore-Fay, Scott (2010). «Cranial osteology and phylogenetic position of the theropod dinosaur Proceratosaurus bradleyi (Woodward, 1910) from the Middle Jurassic of England». Zoological Journal of the Linnean Society 2009 online preprint: p. 155. doi:.
- ↑ Averianov, A. O.; Krasnolutskii, S. A.; and Ivantsov, S. V. (2010). «A new basal coelurosaur (Dinosauria: Theropoda) from the Middle Jurassic of Siberia». Proceedings of the Zoological Institute 314 (1): pp. 42–57.
- ↑ Romer, Alfred S. (1956). Osteology of the Reptiles. Chicago: University of Chicago Press. p. 772pp. ISBN 978-0894649851.
- ↑ Gauthier, Jacques (1986). «Saurischian monophyly and the origin of birds». En Padian, Kevin. (ed.). The Origin of Birds and the Evolution of Flight. Memoirs of the California Academy of Sciences 8. San Francisco, CA: Published by California Academy of Sciences. pp. 1–55. ISBN 978-0940228146.
- ↑ Molnar, Ralph E.; Kurzanov, Sergei M.; & Dong Zhiming. (1990). «Carnosauria». En Weishampel, David B.; Dodson, Peter; & Osmólska, Halszka (eds.).. The Dinosauria (First edición). Berkeley: University of California Press. pp. 169–209. ISBN 978-0520067271.
- ↑ Novas, Fernando E. (1992). «The evolution of the carnivorous dinosaurs». En Sanz, José L.; & Buscalioni, Angela D. (eds.) (en Spanish). Los Dinosaurios y su Entorno Biotico: Actas del Segundo Curso de Paleontología en Cuenca. Cuenca: Instituto "Juan de Valdez". pp. 125–163.
- ↑ Matthew, William D.; & Brown, Barnum. (1922). «The family Deinodontidae, with notice of a new genus from the Cretaceous of Alberta». Bulletin of the American Museum of Natural History 46: pp. 367–385. http://digitallibrary.amnh.org/dspace/handle/2246/1300.
- ↑ von Huene, Friedrich. «Carnivorous Saurischia in Europe since the Triassic». Geological Society of America Bulletin 34 (3): pp. 449–458.
- ↑ Sereno, Paul C. (1999). «The evolution of dinosaurs». Science 284 (5423): pp. 2137–2147. doi:. PMID 10381873.
- ↑ a b c Rauhut, Oliver W.M. (2003). «The interrelationships and evolution of basal theropod dinosaurs». Special Papers in Palaeontology 69: pp. 1–213. ISBN 090170279X.
- ↑ Currie, Philip J.; Hurum, Jørn H; & Sabath, Karol. (2003). «Skull structure and evolution in tyrannosaurid phylogeny» (pdf). Acta Palaeontologica Polonica 48 (2): pp. 227–234. http://app.pan.pl/archive/published/app48/app48-227.pdf.
- ↑ a b Norell, Mark A.; Clark, James M.; Turner, Alan H.; Makovicky, Peter J.; Barsbold, Rinchen; & Rowe, Timothy. (2006). «A new dromaeosaurid theropod from Ukhaa Tolgod (Ömnögov, Mongolia)». American Museum Novitates 3545: pp. 1–51. doi:. http://digitallibrary.amnh.org/dspace/handle/2246/5823.
- ↑ Holtz, Thomas R. (20-09-2005). «RE: Burpee Conference (LONG)». Archives of the Dinosaur Mailing List. Consultado el 18-06-2007.
- ↑ Brusatte, S., Benson, R., Chure, D., Xu, X., Sullivan, C., and Hone, D. (2009). "The first definitive carcharodontosaurid (Dinosauria: Theropoda) from Asia and the delayed ascent of tyrannosaurids." Naturwissenschaften, DOI: 10.1007/s00114-009-0565-2
- ↑ Buffetaut, Eric; Suteethorn, Varavudh; & Tong Haiyan (1996). «The earliest known tyrannosaur from the Lower Cretaceous of Thailand». Nature 381 (6584): pp. 689–691. doi:.
- ↑ a b Holtz, Thomas R.; Molnar, Ralph E.; & Currie, Philip J. (2004). «Basal Tetanurae». En Weishampel, David B.; Dodson, Peter; & Osmólska, Halszka (eds.). The Dinosauria (Second edición). Berkeley: University of California Press. pp. 71–110. ISBN 0-520-24209-2.
- ↑ a b Kirkland, James I.; Britt, Brooks; Burge, Donald L.; Carpenter, Kenneth; Cifelli, Richard; DeCourten, Frank; Eaton, Jeffrey; Hasiotis, Steven; & Lawton, Timothy. (1997). «Lower to Middle Cretaceous Dinosaur faunas of the central Colorado Plateau: a key to understanding 35 million years of tectonics, sedimentology, evolution, and biogeography». Brigham Young University Geology Studies 42 (II): pp. 69–103.
- ↑ Manabe, Makoto (1999). «The early evolution of the Tyrannosauridae in Asia». Journal of Paleontology 73 (6): pp. 1176–1178. http://jpaleontol.geoscienceworld.org/cgi/content/abstract/73/6/1176.
- ↑ Nesov, Lev A. (1995) (en ruso). Dinosaurs of Northern Eurasia: new data about assemblages, ecology and paleobiogeography. St. Petersburg: Scientific Research Institute of the Earth's Crust, St. Petersburg State University. p. 156pp.
- ↑ Fiorillo, Anthony R.; & Gangloff, Roland A. (2000). «Theropod teeth from the Prince Creek Formation (Cretaceous) of northern Alaska, with speculations on Arctic dinosaur paleoecology». Journal of Vertebrate Paleontology 20 (4): pp. 675–682. doi:. http://www.bioone.org/perlserv/?request=get-abstract&doi=10.1671%2F0272-4634%282000%29020%5B0675%3ATTFTPC%5D2.0.CO%3B2.
- ↑ Benson, R. B. J.; Barrett, P. M.; Rich, T. H.; and Vickers-Rich, P. (2010). «A southern tyrant reptile». Science 327 (5973): p. 1613. doi:. PMID 20339066.
- ↑ a b Zhou Zhonghe, Z; Barrett, Paul M.; & Hilton, Jason. (2003). «An exceptionally preserved Lower Cretaceous ecosystem». Nature 421 (6925): pp. 807–814. doi:. PMID 12594504.
- ↑ a b Chen Peiji, Pei-ji; Dong Zhiming; & Zhen Shuonan. (1998). «An exceptionally well-preserved theropod dinosaur from the Yixian Formation of China». Nature 391 (6663): pp. 147–152. doi:.
- ↑ Xu Xing, X; Zhou Zhonghe & Prum, Richard A. (2003). «Branched integumental structures in Sinornithosaurus and the origin of feathers». Nature 410 (6825): pp. 200–204. doi:. PMID 11242078.
- ↑ Lingham-Soliar, Theagarten; Feduccia, Alan & Wang, Xiaolin. (2007). «A new Chinese specimen indicates that ‘protofeathers’ in the Early Cretaceous theropod dinosaur Sinosauropteryx are degraded collagen fibres». Proceedings of the Royal Society of London. Series B, Biological Sciences 270 (1620): pp. 1823–1829. doi:. PMID 17521978.
- ↑ Martin, Larry D.; & Czerkas, Stephan A. (2000). «The fossil record of feather evolution in the Mesozoic». American Zoologist 40 (4): pp. 687–694. doi:. http://www.bioone.org/perlserv/?request=get-abstract&doi=10.1668%2F0003-1569%282000%29040%5B0687%3ATFROFE%5D2.0.CO%3B2.
- ↑ Tanke, D.H. and Brett-Surman, M.K. 2001. Evidence of Hatchling and Nestling-Size Hadrosaurs (Reptilia:Ornithischia) from Dinosaur Provincial Park (Dinosaur Park Formation: Campanian), Alberta, Canada. pp. 206-218. In: Mesozoic Vertebrate Life—New Research Inspired by the Paleontology of Philip J. Currie. Edited by D.H. Tanke and K. Carpenter. Indiana University Press: Bloomington. xviii + 577 pp.
- Weishampel, David. B.; Dodson, Peter; Ósmolska, Halzska. «Tyrannosauroidea». The Dinosauria (2 edición). Berkeley: University of California Press. pp. 880. ISBN 978-0-520-24209-8.
Enlaces externos
Categorías:- Fósiles temporal
- Fósiles
- Tyrannosauroidea


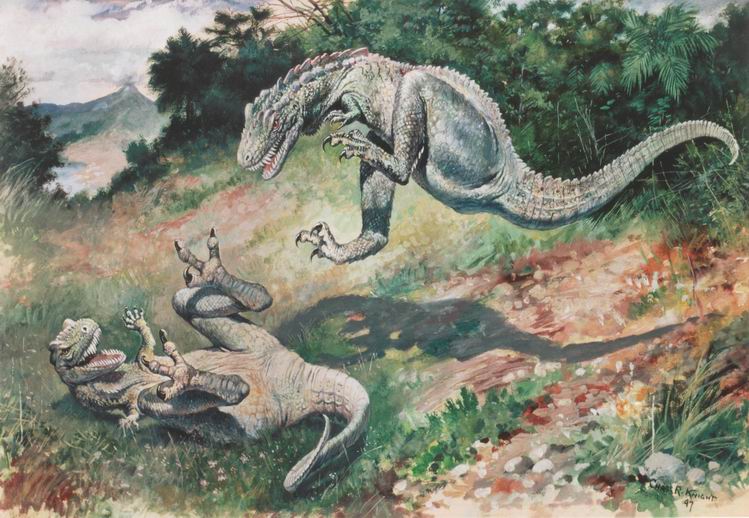

Wikimedia foundation. 2010.