- Agelaius phoeniceus
-
 Tordo sargento
Tordo sargento
Macho exhibiendo su característico plumaje predominantemente negro con la mancha roja en el ala bordeada por la banda amarilla.Estado de conservación 
Preocupación menor (UICN)[1]Clasificación científica Reino: Animalia Filo: Chordata Clase: Aves Orden: Passeriformes Familia: Icteridae Género: Agelaius Especie: A. phoeniceus Nombre binomial Agelaius phoeniceus
(Linnaeus, 1766)[1]Distribución 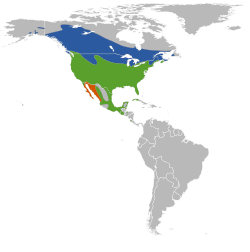
Azul: verano (época de reproducción); verde: durante todo el año; anaranjado: invierno (fuera de la época de reproducción)Subespecies A. p. aciculatus
A. p. arctolegus
A. p. arthuralleni
A. p. brevirostris
A. p. bryanti
A. p. californicus
A. p. caurinus
A. p. floridanus
A. p. fortis
A. p. grinnelli
A. p. gubernator
A. p. littoralis
A. p. mailliardorum
A. p. mearnsi
A. p. megapotamus
A. p. nelsoni
A. p. neutralis
A. p. nevadensis
A. p. nyaritensis
A. p. phoeniceus
A. p. richmondi
A. p. sonoriensis[2]El tordo sargento[2] [3] (Agelaius phoeniceus), también conocido como sargento,[4] tordo alirrojo,[4] tordo capitán[4] o turpial alirrojo,[5] es un ave de hábitos migratorios perteneciente a la familia de los ictéridos, dentro del orden de los paseriformes.[5] Habita en zonas húmedas y terrenos de cultivo de América del Norte y Central.[5] Esta especie presenta un marcado dimorfismo sexual vinculado a una sensible diferencia en su tamaño y al color del plumaje,[6] [7] siendo las hembras principalmente marrones con un intenso veteado ventral y los machos predominantemente negros con un par de manchas rojas bordeadas por una banda amarilla sobre las alas. Los insectos y las semillas constituyen la mayor parte de su dieta.[5] [4] Ha sido considerado una plaga para la agricultura.[6] Se destaca por sus hábitos poligínicos y su alto grado de territorialidad.[6] Quizá sea el ave nativa más abundante de América del norte, con una población estimada de 190 millones de individuos para 1974. Sus poblaciones crecieron enormemente a mediados del siglo XX.[4] Se le considera una especie bajo preocupación menor.[1] [4] Constituye una de las especies de ave más estudiadas de América del norte.
Contenido
Taxonomía
 Agelaius phoeniceus gubernator macho visto desde atrás, mostrando la ausencia de las típicas bandas amarillas debajo de las manchas rojas.
Agelaius phoeniceus gubernator macho visto desde atrás, mostrando la ausencia de las típicas bandas amarillas debajo de las manchas rojas.
El tordo sargento es una de las once especies tradicionalmente clasificadas dentro del género Agelaius y una de las cinco que siguen siendo consideradas parte de éste desde la reclasificación del Congreso Ornitológico Internacional de 2010.[8] Pertenece a la familia de los ictéridos, la cual comprende a aves paseriformes oriundas de América del norte y del sur.[2] [7] Carlos Linneo se refería originalmente a la especie como Oriolus phoeniceus en su obra Systema naturae en el siglo XVIII,[9] pero ésta fue luego reclasificada dentro del género Agelaius junto con otras aves (Vieillot, 1816).[2] [7] [9] [10] Este nombre es una latinización del griego agelaios, “perteneciente a una bandada”. El nombre de la especie, phoeniceus, es una palabra latina que significa “rojo intenso”.[7]
Existen diversas subespecies, las cuales son bastante similares en apariencia.[7] [11] La subespecie típica (A. p. phoeniceus) habita desde Ontario centro-septentrional al norte de Florida y desde el río Mississipi al océano Atlántico.[12] A pesar de las similitudes en la mayor parte de las formas de tordo sargento, en la subespecie de la altiplanicie mexicana, A. p. gubernator, el veteado de la hembra está muy reducido y se restringe a la garganta; el resto del plumaje es marrón muy oscuro.[7] [13] En las subespecies californianas, A. p. californicus y A. p. mailliardorum, el veteado de los ejemplares femeninos también aparece reducido y el plumaje es marrón oscuro, aunque no en el grado de gubernator;[13] asimismo su lista superciliar está ausente o poco desarrollada.[11] Los machos de estas subespecies carecen de la banda amarilla en el ala presente en la mayoría de los miembros masculinos de la especie.[4] [7] [11] El mayito de la ciénaga (Agelaius assimilis), endémico de Cuba, era considerado antiguamente una subespecie de tordo sargento, pero luego se lo reconoció como una especie distinta.[14]
Descripción
 Hembra posada.
Hembra posada. Macho posado.
Macho posado.La coloración del plumaje del macho es completamente negra, a excepción de las plumas cobertoras alares menores (las que cubren los hombros), que son escarlata.[3] En reposo, también exhibe una banda amarilla pálido más pequeña debajo de las manchas rojas,[7] la cual se corresponde con la plumas medianas.[3] Las manchas de los machos de menos de un año de edad, generalmente subordinados, son más pequeñas y anaranjadas que las de los adultos.[15] Estas manchas son vitales en la defensa del territorio.[16] Los machos con las manchas más grandes son más efectivos al ahuyentar a sus rivales no territoriales y tienen mayor éxito en contiendas dentro de aviarios.[17] [18] Al teñirlas de negro como parte de un experimento, el 64% de los machos perdió sus territorios mientras que sólo el 8% de los individuos de control lo hizo. Sin embargo, los machos cuyas alas habían sido teñidas antes de que hubieran conseguido pareja aún podían atraer hembras y reproducirse exitosamente. Aunque las manchas sobre las alas constituyen una señal de amenaza entre los machos, tienen un rol poco importante, si es que acaso tienen alguno, en encuentros intersexuales. Por lo tanto, es probable que las manchas hayan evolucionado en respuesta a las presiones vinculadas a la selección intrasexual.[19] Adicionalmente, ni el tamaño ni la coloración de las mismas están vinculados al éxito reproductivo de los machos respecto de las hembras que no son sus compañeras, es decir, aquellas con las que copulan eventualmente.[16]
Las partes superiores de la hembra son de color marrón, mientras que las inferiores están cubiertas por un intenso veteado blanco y oscuro;[5] presenta asimismo una lista superciliar blanquecina.[4] Las hembras de un año exhiben una mancha rosada o salmón sobre los hombros y una coloración rosada claro en el rostro y debajo de este, mientras que las de mayor edad muestran una mancha más carmesí sobre los hombros y una tonalidad rosada oscuro en el rostro y debajo del mismo.[20] Los juveniles son similares a las hembras adultas, pero presentan una coloración abdominal más pálida.[7]
 Hembra posada sobre un alambre.
Hembra posada sobre un alambre.La envergadura de esta especie es de entre 31 y 40 cm aproximadamente.[4] El pico en ambos sexos es largo, delgado y puntiagudo.[5] [21] Tanto el pico del macho como las patas y los ojos son negros;[5] en la hembra el pico es marrón oscuro en la mitad superior y claro en la inferior.[22] La cola es de longitud media y redondeada.[4] [7] Así como en otras especies poligínicas, existe en el tordo sargento un considerable dimorfismo sexual tanto en el plumaje como en el tamaño, pesando los machos entre 65 y 80 g y las hembras alrededor de 35 g.[6] [23] Los machos son un 50% más pesados que las hembras[17] y un 20% más grandes que estas respecto de la longitud de sus alas.[24] La tendencia hacia un mayor dimorfismo en el tamaño de las especies de ictéridos no monógamas indica que el mayor tamaño de los machos ha evolucionado a causa de la selección sexual.[24]
El macho es inconfundible, excepto en California y algunas áreas en sus alrededores, en donde cohabita con el tordo tricolor (Agelaius tricolor), de aspecto similar.[4] [7] Sin embargo, esa especie presenta manchas de un color rojo más intenso, rodeadas de blanco en vez de amarillo.[4] [7] En vuelo, cuando la coloración del plumaje no es fácilmente distinguible, aún es posible discriminar entre estos e ictéridos menos emparentados —tales como el zanate norteño (Quiscalus quiscula) y el tordo cabecicafé (Molothrus ater)— por su particular silueta y su vuelo ondulante.[7]
Distribución, hábitat y migración
 Macho posado sobre una conífera.
Macho posado sobre una conífera.El tordo sargento se reproduce desde Alaska centro-oriental y Yukón hasta el norte de Costa Rica y del Atlántico al Pacífico.[6] Los ejemplares que habitan en la región norte son migratorios y pasan el invierno en el sur de Estados Unidos y en Centroamérica,[22] aunque muchos lo hacen tan al norte como el sur de Canadá.[6] Las migraciones comienzan en septiembre u octubre, aunque ocasionalmente lo hacen en agosto.[22] En el oeste de América del norte, así como en latitudes un tanto más australes hacia el este, las poblaciones son generalmente sedentarias.[22]
Esta especie habita mayormente en llanuras. Por lo general, prefiere los humedales y puede hallársele en pantanos, particularmente donde crezcan totoras (Typha spp.).[7] En gran parte de su área de distribución, constituye el ave paseriforme más abundante en los pantanos en los que se reproduce.[6] También se encuentra presente en áreas sin mucha agua, en donde habita en campo abierto (a menudo áreas agrícolas) y bosques caducifolios poco densos.[22] Está muy diseminado por toda América del norte, excepto en el desierto árido, altas cadenas montañosas y regiones árticas o de forestación densa.[25]
En un estudio en Illinois central, las poblaciones de tordos sargento eran mucho mayores a lo largo de carreteras interestatales que a los lados de rutas de menor envergadura. Esto probablemente se deba a que las primeras se ubicaban en medio de amplias franjas dominadas por el pasto azul de Kentucky (Poa pratensis), a menudo en compañía de arbustos y pequeños árboles. Por otro lado, las pequeñas dimensiones del área no cultivada a los lados de las rutas de menor envergadura probablemente brindaran una cantidad más reducida de sustratos adecuados para la nidificación.[26]
Comportamiento
 Hembra posada sobre un tronco.
Hembra posada sobre un tronco.Se trata de aves territoriales, poligínicas, gregarias y migratorias de corta distancia. El comportamiento de los machos hace que su presencia sea fácilmente percibida: se posan en lugares altos y cantan durante todo el día. Las hembras suelen quedarse en lo bajo, merodeando entre la vegetación y construyendo sus extraordinarios nidos. Puede encontrárseles en jardines hogaareños, particularmente durante su migración, si se ha esparcido semillas sobre el suelo. También se les puede ver posados sobre cables de teléfono. Descansan en bandadas durante todo el año. En verano, duermen en pequeñas cantidades en los humedales donde se reproducen. En invierno, se congregan en grandes bandadas de hasta varios millones de individuos de varias especies. Cada mañana, estas bandadas masivas se disgregan y se alejan hasta 80 km para alimentarse; luego se reagrupan en la noche.[4] En los sitios de descanso comunal, los machos de mayor edad son dominantes respecto de los que han eclosionado ese mismo año y ocupan lugares centrales en la vegetación densa, en donde el riesgo de depredación es menor.[27] Su manera de volar es característica, con raptos de rápido batimiento de alas, entre los cuales se producen pequeñas pausas intermitentes.[25]
Durante la temporada reproductiva, los machos a menudo vigilan los nidos activos desde puntos de observación prominentes. En un estudio experimental, los machos pasaban significativamente más tiempo dentro de los 10 m desde sus nidos tras la presentación de un depredador simulado que en otros momentos del mismo día, y muchos de ellos revisaban sus nidos inmediatamene después de que aquel fuera removido.[28]
Durante la época de reproducción, tanto machos como hembras suelen alimentarse fuera de sus territorios.[6] A finales del verano y en otoño, se les encuentra alimentándose en campo abierto junto a otros ictéridos —comúnmente zanates norteños (Quiscalus mexicanus) y tordos cabecicafé (Molothrus ater)— y estorninos pintos (Sturnus vulgaris) en bandadas que alcanzan los miles o millones de individuos.[4] [7] [25] [29] También se congregan con tordos canadienses (Euphagus carolinus).[30]
Vocalizaciones
Entre las especies con un área de distribución templada, es relativamente raro que la hembra cante. El tordo sargento es una de las pocas aves conocidas de zonas templadas en las que machos y hembras cantan canciones distintas.[12] La variedad del repertorio de los machos guarda relación con la edad de los individuos, su experiencia reproductiva y el tamaño de sus harenes.[31] Los machos con repertorios más grandes son más experimentados y consiguen un mayor número de compañeras respecto de los que exhiben repertorios pequeños.[32] Los machos tienen repertorios de entre dos y ocho canciones,[33] y a menudo cambian de canción cuando se posan en un nuevo sitio.[34] Asimismo, cambian de canción más frecuentemente y emiten más tipos de canciones en presencia de una hembra que de otro macho. Esto respalda la hipótesis de que los repertorios de los tordos sargento machos evolucionaron principalmente como respuesta a la selección intersexual.[33] [31] No obstante ello, la evidencia experimental sugiere que los repertorios de los machos inciden en su éxito tanto en la atracción de una pareja como en contiendas entre individuos de su mismo sexo.[33] Una variedad más amplia de canciones parece suscitar respuestas de cortejo en las hembras con mayor eficacia y favorecer la defensa del territorio respecto de la intrusión de competidores.[33] La presentación del repertorio es más efectiva para ahuyentar a los intrusos que la repetición de una única canción.[31] [34]
Por otro lado, la cantidad de canciones emitidas por los machos por minuto parece no guardar relación con el tamaño de sus harenes. Asimismo, la cantidad de canciones emitidas no varía significativamente entre los machos territoriales y los machos originalmente no territoriales que reemplazan a anteriores propietarios de un territorio que hayan desaparecido; sin embargo, al menos respecto de estos últimos machos, disminuye a medida que transcurre el tiempo desde el momento en que obtienen posesión del territorio, lo cual podría estar relacionado con el hecho de que los machos no territoriales compiten más intensamente por un territorio abandonado poco después de la desaparición de su dueño original. La cantidad de canciones tampoco difiere entre los machos cuyos territorios están aislados de los de otros machos y aquellos cuyos territorios son contiguos a otros, y entre estos últimos tampoco depende del número de vecinos con los que comparta límites territoriales. En cambio, sí aumenta ante la intrusión de un potencial competidor. La evidencia indica, por lo tanto, que la cantidad de canciones emitidas no se ve modificada en respuesta a interacciones pasivas, pero se incrementa dramáticamente cuando la interacción se vuelve más amenazante.[31]
Cuando se suministra alimento experimentalmente, los machos de muchas especies, incluyendo el tordo sargento, aumentan la cantidad de canciones emitidas, lo cual sugiere que esta podría señalar la calidad del territorio. Es posible que las hembras interpreten esas señales puesto que los territorios de tordo sargento con alimento adicional atraen harenes más numerosos y las hembras en esos territorios se reproducen más temprano. Sin embargo, la añadidura de alimento podría resultar en una variación en la cantidad de canciones superior a la que se produciría en circunstancias naturales. En favor de esta teoría, cabe destacar que un estudio no logró detectar una relación entre la intensidad de las exhibiciones de los machos y el tamaño de los harenes en experimentos en los que no se proveyó de aliemento adicional. La tendencia de los machos a alimentarse fuera de sus territorios también poría minimizar cualquier correspondencia entre la abundancia de alimento dentro de sus territorios y la cantidad de canciones emitidas. En síntesis, la evidencia sugiere que la disponibilidad de alimento en situaciones naturales no varía lo suficiente como para afectar la cantidad de canciones de los machos de tal manera que esta influya a su vez sobre la decisión de las hembras respecto del territorio en el que se establecen.[31]
Las hembras reproductivamente exitosas, es decir aquellas cuyos nidos albergan al menos un polluelo que logra independizarse, tienden a responder más frecuentemente a las canciones de sus compañeros con el llamado "chit" desde sus nidos que las hembras no exitosas. Estos llamados parecen proporcionar un beneficio a las hembras de tordo sargento al mantener la vigilancia de los machos respecto de los depredadores.[35] El llamado "chit" es emitido con relativa consistencia a lo largo del ciclo de nidificación, pero las hembras rara vez dan estos llamados una vez que sus polluelos abandonan el nido. Las hembras que no reciben asistencia por parte del macho en la alimentación de sus polluelos emiten el llamado más frecuentemente que las que son asistidas.[36]
Alimentación
 Plantas de Panicum virgatum, de cuyas semillas se alimenta el tordo sargento.
Plantas de Panicum virgatum, de cuyas semillas se alimenta el tordo sargento.Esta especie es omnívora.[7] [22] Se alimenta principalmente de productos vegetales —particularmente en otoño e invierno—,[5] tales como semillas de hierbas —ambrosías (Ambrosia spp.), arrancamoños (Xanthium spp.), etc.—[4] y granos (maíz, arroz, trigo, girasol, etc.),[4] pero alrededor de un cuarto de su alimentación —y significativamente más durante la temporada de reproducción— consiste en insectos y otros animales pequeños.[7] [22] Consume mayormente insectos como libélulas, caballitos del diablo, mariposas, polillas, moscas, pero también caracoles, ranas, huevos, carroña, lombrices y arácnidos.[7] El tordo sargento consigue sus presas entre las plantas o atrapándolas en vuelo.[7] [22] A veces las obtiene explorando la base de plantas acuáticas con su pico delgado, abriendo orificios para llegar a los insectos escondidos adentro.[4] [22] En temporada, come además arándanos, mora y otras frutas.[7]
 Plantas de Xerophyllum tenax, de cuyas semillas se alimenta el tordo sargento.
Plantas de Xerophyllum tenax, de cuyas semillas se alimenta el tordo sargento.Un estudio llevado a cabo en 1900 examinando los contenidos estomacales de 1.083 tordos obtenidos a lo largo de todos los meses del año en la mayor parte de su área de distribución en Estados Unidos y Canadá reveló que estas aves prefieren las semillas de hierbas e insectos a los granos de cultivo. El estudio indicó que el 73,4% de su dieta es de origen vegetal, frente a un 26,6% de origen animal. A continuación se citan los porcentajes con mayor especificidad: escarabajos predadores 2,5%, gorgojos (curculiónidos) 4,1%, otros escarabajos 3,5%, orugas 5,9%, saltamontes 4,7%, otros insectos 4,1%, arañas y miriápodos 1,3%, otras presas 0,5%, frutas 0,6%, maíz 4,6%, avena 6,3%, trigo 2,2%, otros granos 0,8%, semillas de hierbas —incluyendo Panicum spp., Xerophyllum tenax, Ambrosia spp. y Polygonum spp.— 54,6% y otros alimentos de origen vegetal 4,3%. En el mes de noviembre el consumo de semillas de hierbas representa hasta un 97% de su alimentación. La ingesta de frutas, tales como moras y arándanos, es poco usual. De los 84 especímenes examinados por el científico Franklin Hiram King en Wisconsin, 37 habían ingerido semillas de hierbas y maíz conjuntamente, 31 sólo semillas, 7 sólo maíz, 3 centeno, 2 avena, 8 trigo y 2 hierba tierna; 5 habían comido un total de siete escarabajos, 4 siete saltamontes, 1 una polilla y 1 una oruga; 8 habían comido pequeños moluscos.[25] Una investigación realizada entre noviembre y mediados de marzo en el oeste de Tennessee dio como resultado que su alimentación en esos meses estaba compuesta de maíz en un 30% y de semillas de 15 géneros de hierbas en un 60%.[29]
En el suroeste de Quebec las poblaciones de tordos sargento se duplicaron con creces entre 1966 y 1981, aparentemente en respuesta al desarrollo de la producción de maíz. La mayor disponibilidad de granos residuales en primavera y verano (la temporada reproductiva) probablemente jugara un rol fundamental en el crecimiento de la población de tordos sargento.[37]
Reproducción
 Macho echándose a volar.
Macho echándose a volar.Importancia de la simetría y del estatus parasitológico
A diferencia de lo que sucede en otras especies, no se encontró evidencia de que sutiles imperfecciones en la simetría bilateral de los machos de tordo sargento tuviera relación con un peor estado de salud o menores posibilidades de sobrevivir fuera de la temporada reproductiva,[38] [39] un menor éxito al competir por territorios, compañeras u otras hembras con las que copular que no fueran sus compañeras,[40] [39] o un menor éxito en la competencia intrasexual por el alimento.[41] [39] Debido a que los resultados de estudios sobre la simetría bilateral de los tordos sargento machos no son consistentes con la premisa de que un alto grado de asimetría indicaría una calidad fenotípica o genotípica inferior, quizá no sorprenda que las hembras de esta especie no parezcan tomar la simetría como criterio en la elección de pareja.[40]
Los machos con parásitos en la sangre tienen las mismas posibilidades de adquirir un territorio y de sobrevivir de un año al otro que los no parasitados. De manera similar, las hembras parasitadas y no parasitadas no muestran diferencias en referencia a cuán temprano comienzan a anidar, la cantidad de huevos que depositan y sus posibilidades de supervivencia de un año al siguiente. Por otro lado, los rasgos sexuales secundarios masculinos, particularmente la agresión intrasexual, revelaban el estatus parasitológico bastante acertadamente. El plumaje y los rasgos morfológicos también permiten discriminar entre individuos con y sin parásitos entre las hembras. Sin embargo, los patrones de apareamiento aparentemente no guardan relación con el estatus parasitológico de los machos o de las hembras.[42]
Territorialismo y cortejo
 Macho cantando.
Macho cantando.La época de reproducción se extiende entre abril y julio.[40] [43] Los machos llegan a las áreas de reproducción unas tres semanas antes que las hembras para establecer sus territorios;[44] luego las hembras se asientan dentro de los mismos.[45] Normalmente, los machos se hacen de uno por primera vez a los dos o tres años de edad:[40] no suelen reproducirse en su primer año.[46] [47] Las hembras, en cambio, se reproducen ya en la primera temporada reproductiva posterior a su nacimiento.[48] Entre los individuos de más de dos años, los machos no territoriales mayores no son más exitosos que los más jóvenes respecto de la eventual adquisición de un territorio.[49] En ocasiones, establecen sus territorios en estanques de castores (Castor canadensis).[40] [43] Los territorios pueden hallarse tanto aislados como contiguos a otros.[17] [49] Los límites de los territorios están bien definidos y se mantienen a lo largo de la temporada reproductiva.[50] Es raro que un macho cambie de territorio durante la época de reproducción.[49] Los machos no territoriales, por otro lado, se deplazan sobre amplias áreas geográficas.[49]
El establecimiento de un territorio es de gran importancia para el éxito reproductivo de los machos puesto que los machos no territoriales representan colectivamente menos del 5% de las fecundaciones. Debido a que entre un 35 y un 80% de los machos reproductivamente competentes no logran conseguir un territorio cada año, la competencia por estos es intensa.[40] La adquisición de un territorio puede implicar el desplazamiento de un residente ya establecido.[44] [51] El éxito reproductivo de los tordos sargento machos también varía en función de la calidad de los territorios. Aquellos que se establecen en pantanos tienden a engendrar una descendencia más numerosa que los que se asientan en campos en tierras altas, en gran parte debido a que los machos con territorios pantanosos típicamente atraen a más compañeras.[40]
El tordo sargento puede ser muy agresivo al defender su territorio de la intrusión de otras aves:[4] ataca a menudo a potenciales competidores de su especie, tordos tricolores (Agelaius tricolor) y tordos cabeciamarillos (Xanthocephalus xanthocephalus),[22] e incluso aves de mucho mayor tamaño como cuervos, urracas (Pica pica), aves de presa —halcones y águilas pescadoras (Pandion haliaetus), por ejemplo—[25] y garzas.[7] Otras especies contra las que se demuestra persistentemente agresivo incluyen el zanate norteño (Quiscalus quiscula) y el chivirín pantanero (Cistothorus palustris).[50] Es bien conocida la predisposición del tordo sargento macho a atacar a seres humanos durante la temporada reproductiva, aunque los encuentros deliberados con cernícalos americanos (Falco sparverius) son probablemente poco comunes; no obstante ello, se sabe de triunfos de tordos sargento machos en enfrentamientos pico a pico en medio del aire con individuos de esa especie.[50] En ocasiones, defienden nidos de sus vecinos cuando han engendrado polluelos en ellos.[43] [52]
La importancia de las peleas parece ser limitada en la defensa del territorio respecto de otros machos. Una vez que un territorio es ocupado, es raro que su dueño necesite ser agresivo para defenderlo de machos no territoriales debido a que la mayoría de estos espera a que el dueño de un territorio desaparezca para establecerse como macho territorial en lugar de hacer uso de la agresión para reclamar uno. Los territorios abandonados a menudo vuelven a ser ocupados dentro de unos minutos (en alrededor de un 10% de los casos, son anexionados al territorio de algún macho territorial vecino). Esto sugiere que los machos no territoriales que eventualmente logran adquirir un territorio son simplemente los primeros en descubrir un espacio dejado en desuso.[49]
En un estudio, más de la mitad de los machos marcados con bandas rojas perdió su territorio, mientras que todos los que habían sido identificados con bandas negras retuvieron los suyos. Asimismo, los primeros sufrieron una cantidad de intrusiones mucho mayor en sus respectivos territorios que los segundos.[53]
El cortejo incluye vocalizaciones en una posición inclinada y batimientos rápidos y notorios de las alas por parte de los ejemplares masculinos.[22] Estos inflan su plumaje, levantan sus hombros y despliegan sus colas mientras cantan.[22] [25] A medida que sus demostraciones aumentan en intensidad, arquean más las alas dejando ver las manchas rojas en sus hombros.[22] Las hembras responden con una vocalización e inclinación similares.[22]
Para el momento en que se producen las cópulas, disminuyen significativamente la cantidad de canciones y exhibiciones de los machos y el número de intrusiones a territorios ajenos. Antes del apareamiento muchos aspectos del territorialismo llegan a su punto máximo. Después de este, la frecuencia de muchas de las conductas territoriales decrece y los machos territoriales se ocupan principalmente de la defensa de las hembras, los huevos y los polluelos de la depredación. Experimentos de remoción sistemática de las aves de sus territorios sugieren que la población extra de machos que se encuentra presente en los pantanos antes de las cópulas desaparece después de las mismas.[54]
Filopatría
Se trata de una especie filopátrica, es decir, regresa a reproducirse a las mismas áreas año tras año. Algunos machos pueden regresar a los mismos territorios hasta por 8 o 9 años. En el este de Ontario, en un período de seis años el 55,3% de los machos regresó al área de estudio en al menos una oportunidad; el 4,4% de esos machos cambió de pantano entre años, el 14,7% cambió de territorio dentro del mismo pantano y el 80,9% volvió a ocupar el mismo territorio. El 41,3% de las hembras se reprodujo dentro del área de estudio durante más de un año; el 75,7% de esas hembras anidó en el mismo pantano que el año anterior. Cuando las hembras tenían la oportunidad de anidar en el territorio del mismo macho que el año anterior —es decir, cuando tanto el macho como la hembra habían vuelto al mismo pantano—, el 58,7% de las hembras lo hizo. Sin embargo, las hembras que regresan a los mismos pantanos parecen ser más fieles a áreas generales dentro de estos que a machos en particular.[43] La marcada tendencia de las hembras a anidar en las mismas áreas en años consecutivos repercute significativamente en el éxito de los machos para encontrar pareja debido a que los nuevos machos “heredan” los harenes de los anteriores dueños de determinados territorios.[55]
En otro estudio, alrededor del 70% de los machos que se reprodujeron en más de una temporada retuvieron sus territorios originales. La mayor parte del 30% de los machos que cambiaron de territorio se estableció a menos de 200 m de su territorio original, a menudo en uno adyacente. Los machos que se habían desplazado menos de 200 m tendían a mejorar su éxito reproductivo, lo que respalda la hipótesis de que estos sólo cambiarían de territorio voluntariamente si esperasen mejorar sus posibilidades de reproducirse de manera significativa; por otro lado, los machos que se desplazaban grandes distancias no presentaban incrementos en su éxito reproductivo, de modo que estos movimientos quizá hayan sido involuntarios.[56]
Vínculo intersexual
Los tordos sargento son poligínicos y los machos territoriales pueden alcanzar las 15 compañeras en algunos casos,[57] aunque normalmente tienen dos o tres.[17] [49] El tamaño de los harenes suele ser mayor en los pantanos que en hábitats de tierras altas; además, pareciera ser más variable en pantanos que en otros hábitats.[16] Tanto la calidad del macho como la de su territorio influyen en grados variables sobre el éxito reproductivo femenino y, en consecuencia, sobre la elección de pareja por parte de las hembras.[58] Un estudio determinó que los machos más grandes engendraban una mayor cantidad de polluelos dentro de sus territorios, principalmente debido a que conseguían harenes más numerosos, y que el éxito de los machos de mayor tamaño no parecía ser consecuencia de que estos tuvieran territorios de mayor calidad o extensión.[59] Adicionalmente, son más las hembras que anidan en territorios de machos experimentados que en los de machos jóvenes.[43] Aunque el éxito reproductivo de las hembras que componen el harén de determinado macho parece no estar vinculado a la edad de éste, aparentemente guarda una relación positiva con su tamaño.[40] Por otro lado, existe un consenso general sobre la importancia de la calidad del territorio respecto de la elección de pareja por parte de la hembra.[60] Investigaciones realizadas en Washington, por ejemplo, arrojaron como resultado que la elección del sitio de nidificación está más vinculada a la calidad del territorio que a la del macho.[61] [24]
Asimismo, las hembras frecuentemente copulan con otros machos.[4] En consecuencia, el compañero de la hembra engendra sólo entre dos tercios y tres cuartos de los polluelos que eclosionan en su territorio,[45] existiendo nidadas mixtas en las que los polluelos de un mismo nido fueron concebidos por distintos padres.[7] Un estudio encontró que mientras que el 21% de los polluelos muestreados no había sido engendrado por el dueño del territorio en el que había nacido, el 36% de las nidadas contenía al menos un polluelo ilegítimo;[38] en otro, casi la mitad de los nidos albergaba al menos un polluelo hijo de otro padre.[62] La mayor parte de los polluelos engendrados por machos que no son los compañeros de sus madres (el 77% de ellos en un estudio en el este de Ontario)[40] son hijos de algún otro que defienda un territorio adyacente.[40] [62] [63] Los machos reproductivamente más exitosos con respecto a las hembras que no son parejas suyas también lo son dentro de su harén, de modo que las cópulas casuales con otras hembras no afectan su éxito reproductivo dentro de los límites de sus territorios.[64]
La mayoría de las cópulas entre individuos que no constituyen una pareja (el 78% de ellas en un estudio de una población occidental) se produce fuera del territorio de nidificación de la hembra en donde las parejas copulan típicamente. Las hembras que anidan en áreas con un gran número de machos producen una cantidad significativamente mayor de polluelos hijos de otro padre que las que anidan pantanos con pocos vecinos machos. Las estrategias de los machos para proteger su paternidad no son muy efectivas y se limitan primordialmente a vigilar los límites de sus territorios y en menor medida a impedir cópulas de sus hembras con otros machos fuera del pantano cuando estas son visibles.[65]
Aunque algunos machos no engendran polluelos con hembras que no forman parte de su harén, estas cópulas con hembras que no constituyen parejas suyas contribuyen significativamente a incrementar el éxito reproductivo de otros[40] (en promedio, representan más del 20% del éxito reproductivo de cada macho).[62] En el este de Ontario, por ejemplo, se descubrió un macho territorial que había engendrado 15 polluelos con estas hembras, distribuidos en seis nidadas criadas en los territorios de cuatro machos distintos.[40] Los machos de mayor edad muestran una marcada tendencia a ser más exitosos respecto de la reproducción fuera de su harén.[40] [59]
Nido y huevos
Esta especie anida en colonias un tanto disgregadas, en parte debido a que el hábitat pantanoso adecuado para ello es escaso.[4] Típicamente el nido es ubicado cerca del suelo —o de la superficie del agua en áreas pantanosas—, entre totoras (Typha spp.),[25] [40] ciperáceas —por ejemplo, de los géneros Scirpus y Carex—[25] y carrizos (Phragmites australis) en humedales; en varas de oro (Solidago spp.),[25] moreras (Morus spp.), sauces (Salix spp.)[25] y alisos (Alnus spp.)[25] en tierras más altas; o en plantas de trigo, cebada, alfalfa[25] y arroz.[4] También puede hallárseles en ciruelos americanos (Prunus americana).[66] Suelen estar situado a una altura de entre 0,3 y 1,5 m encima del agua en los humedales.[3] Otras fuentes extienden esas cifras a entre 7,6 cm y 4,3 metros.[7] También se ha observado la nidificación sobre el suelo en campo abierto y tierras de labor.[5] [25] La densidad de los nidos es mayor en pantanos de totoras que en campos en tierras altas.[31] Los nidos correspondientes a las compañeras de un mismo macho iniciados con poca diferencia de tiempo no necesariamente son los más próximos entre sí en el espacio.[57] Las hembras de sólo un año de edad comienzan a anidar considerablemente más tarde en la temporada reproductiva que las de mayor edad.[67]
La confección del nido, llevada a cabo por la hembra y completada en alrededor de entre tres y seis días,[7] [25] es realizada entre marzo y mayo; usualmente, cuanto más al sur se vaya, antes comienza el proceso de construcción.[22] Exteriormente, está compuesto por una considerable cantidad de hojas de cárices (Carex spp.) o de otras plantas recolectadas en lugares húmedos y ocasionalmente por hojas de Iris, atadas a las hierbas o tallos circundantes a modo de soporte; en casos en los que el nido se halla directamente sobre el suelo esta precaución de fijarlo en su lugar es dejada de lado.[25] En la parte interna, se coloca madera en descomposición, césped, turba o barro, todo esto revestido con pequeños tallos secos de césped.[25] En la década de 1930, se descubrió un nido sujeto a 18 tallos que contenía 34 tiras de corteza de sauce de hasta alrededor de 86 cm, 142 hojas de totoras de hasta unos 53 cm y 705 trocitos de césped.[4] [25] Cuando está terminado, el nido, en forma de taza, mide entre 10 y 18 cm de ancho por entre 8 y 18 cm de profundidad aproximadamente.[4]
En gran parte de su área de distribución, las puestas constan comúnmente de tres o cuatro huevos.[68] Presentan una coloración verde azulada pálida, con machas marrones, púrpuras o negras, la mayoría de las cuales dispuesta alrededor del extremo más ancho.[7] [22] La medición de 380 huevos de la colección del Museo Nacional de Estados Unidos dio un promedio de 24,80 por 17,55 mm, siendo el más grande de 27,94 por 19,05 mm y el más chico de 20,57 por 15,75 mm.[25]
Las parejas crían habitualmente dos y ocasionalmente tres nidadas por temporada, cada una de las cuales en un nuevo nido.[7] [21] El segundo es construido en las cercanías del primero, frecuentemente dentro de los 3 metros de distancia de aquél, incluso si éste ha sido atacado o destruido.[25] A menudo la construcción del segundo nido comienza antes de que la primera nidada haya abandonado el suyo.[25] Algunas hembras de tordo sargento pueden anidar hasta cuatro o cinco veces en una temporada si sus nidadas perecen antes de independizarse.[55]
Crianza de los polluelos
 El macho colabora en la crianza.
El macho colabora en la crianza.El período de incubación ronda los 11 días.[31] Los polluelos de una misma nidada tienden a romper el cascarón en diferentes momentos.[22] Diversos estudios sobre el porcentaje que representan los pequeños de cada sexo sobre el total de los polluelos en los que se utilizaron el peso y la longitud del tarso como métodos de identificación del sexo en individuos cuya estadía en el nido ya era avanzada dieron como resultado una menor cantidad de machos que de hembras, aunque las proporciones no eran significativamente distintas del 50%.[68]
Los pequeños son alimentados generalmente con un 100% de materia animal; su dieta está compuesta de preponderantemente de insectos con arácnicos y moluscos ocasionales así como otros tipos de alimentos.[69] Una fracción del alimento dado a los polluelos se extrae de las cercanías del nido, pero gran parte de este es recolectado en áreas fuera del territorio del macho y en muchos casos incluso fuera del hábitat de nidificación a bastante distancia del nido.[69] Los padres comienzan a alimentarlos con porciones pequeñas que aumentan progresivamente.[22] Las hembras son quienes más se ocupan de esta tarea, aunque en ocasiones el macho también participa.[22] [60] [68] [69] Durante el período que se extiende entre el abandono del nido por parte de los polluelos y la adquisición de la independencia respecto de sus padres, el macho los alimenta regularmente.[69] El desarrollo de los polluelos y su peso cuando abandonan el nido son similares entre los que son criados en pantanos y aquellos que crecen en tierras altas.[70] [47]
Las hembras de menor edad tienen nidadas más pequeñas que las más experimentadas.[67] En las nidadas de tamaño medio, el sexo de los compañeros de nido no parece afectar el desarrollo de cada polluelo: así, por ejemplo, un individuo de cualquier sexo tiene las mismas probabilidades de encontrarse en buen estado si sus tres hermanos son machos o hembras.[68] El éxito reproductivo tanto de las nidadas naturalmente numerosas como de aquellas experimentalmente agrandadas parece ser reducido, presumiblemente debido a que la satisfacción de sus necesidades pondría a los padres bajo una presión mayor a la ideal.[68]
Desarrollo de los polluelos
Se trata de una especie altricial:[71] [72] al nacer, los polluelos no pueden ver y están desprovistos de plumas.[4] [21] Al eclosionar, los polluelos de distintos sexos pesan prácticamente lo mismo.[68] Hacia su octavo día de vida, sin embargo, casi todos los machos exceden los 33 g, mientras que casi todas las hembras pesan menos de 33 g,[67] lo cual puede utilizarse a modo de criterio para la identificación del sexo de los polluelos,[67] [47] puesto que para entonces ya no hay virtualmente superposiciones entre los pesos de los machos y los de las hembras.[68] Las hembras tienden a abandonar el nido ligeramente antes que los machos, lo cual se evidencia en el hecho de que, entre los polluelos aún presentes en el nido o lo suficientemente cerca de este como para ser capturados y pesados unos diez u once días después de la eclosión, la proporción de machos es significativamente más elevada que en días anteriores.[68] Para el momento en que aprenden a volar, el peso de los individuos de ambos sexos representa aproximadamente dos tercios del de un adulto; ganan la mayor parte del peso adicional durante el período posterior al abandono del nido en el que aún dependen de los cuidados de sus padres.[68] Aunque los machos maduran sexualmente en la primera temporada reproductiva tras abandonar el nido (es decir, aproximadamente al año de edad), no adquieren su plumaje adulto completo sino hasta al año siguiente.[49]
La masa de los dos músculos pectorales en los tordos sargento representa un 1,8% de la masa corporal total al nacer, pero crece en mayor proporción qeue el resto del cuerpo durante el período postnatal: alcanza el 5,1% a los 8 días y el 6,6% a los 10 días de nacidos. Tras abandonar el nido se produce un incremento más significativo, llegando a representar los dos músculos pectorales el 15,5% de la masa corporal total de los adultos.[71]
Desarrollo de la capacidad termogénica
Permanecen relativamente poco tiempo en el nido, unos 10 o 12 días, tiempo en el cual su peso inicial de 3,5 g tras la eclosión se multiplica entre ocho y once veces.[72] Su capacidad termogénica mejora marcadamente durante el mismo período, transformando a los polluelos de pasivos ectotermos antes de los 3 días de edad en competentes endodermos para el momento en el que dejan el nido.[71] [72] A la edad de 3 días, el metabolismo de los polluelos disminuye sustancialmente en respuesta a caídas graduales en la temperatura.[71] Las mejorías más abruptas en la habilidad para regular la temperatura corporal en esta especie se producen entre el quinto y el octavo día de vida: los polluelos de 8 días pueden mantener su temperatura corporal en un rango mucho mayor de temperaturas ambiente que los de 5 días.[71] [72]
Entre los 2 y los 6 días de edad, las nidadas tomadas en su conjunto logran mantener temperaturas corporales muy superiores con respecto a los individuos; por ejemplo, las nidadas de 6 días generalmente mantienen las temperaturas corporales por encima de los 35 °C incluso a una temperatura ambiente de 5 °C, mientras que las temperaturas corporales de polluelos de esa misma edad tomados individualmente caen lo suficiente como para acercarse a la temperatura ambiente ante la exposición a una temperatura tal de 15 °C y descienden por debajo de los 35 °C incluso a una temperatura ambiente de 25 °C. Los individuos de 8 días logran regular su temperatura corporal a temperaturas ambiente de 15 °C y a veces incluso de 5 °C, pero el costo metabólico que esto representa es proporcionalmente alto respecto de aquel en el que incurriría una nidada. La termogénesis no es controlada por la termorregulación sino hasta los 4 o 5 días de edad en los tordos sargento, aunque las nidadas pueden modular su aislamiento térmico homeostáticamente al menos desde los 3 días. Esto se debe a que los polluelos en el nido experimentan un aislamiento térmico superior al de polluelos aislados gracias a la protección del nido mismo y a la posibilidad de acurrucarse junto a sus pares y así el conjunto de los hermanos logra combinar sus capacidades termogénicas.[73]
Se reconocen dos fases en el desarrollo de estos polluelos altriciales. En la primera fase (denominada “fase inercial”), aunque la termogénesis no se encuentra bajo el control termorregulatorio, las temperaturas corporales de las nidadas en el nido pueden permanecer bastante por encima de la temperatura ambiente durante los largos períodos de ausencia parental en virtud de la inercia térmica. En la fase dos (llamada “fase termorregulatoria”), las nidadas han adquirido la capacidad de termorregularse fisiológicamente, lo que implica una estabilización más efectiva de las temperaturas corporales y costos metabólicos más elevados que en la fase inercial. En los tordos sargento, al menos entre un 53% y un 72% del incremento del contenido de energía corporal resultante del crecimiento entre la eclosión y el momento del abandono del nido se produce durante la fase termorregulatoria.[73]
Factores influyentes sobre la reproducción
En la mayoría de las colonias tanto grandes como pequeñas, el porcentaje de nidos atacados por depredadores es menor durante el apogeo de la actividad vinculada a la anidación que más tarde.[74] En un estudio en el condado de Grant (Washington), menos del 30% de los nidos no parasitados de fines de la época de reproducción fue exitoso, mientras que hasta el 60% de los nidos no parasitados de comienzos de la temporada acogieron polluelos que lograron independizarse.[75]
Las poblaciones que anidan en pantanos tienen mayor éxito reproductivo que las que lo hacen en tierras altas debido a que en el primer caso es menor la proporción de nidos destruidos por depredadores.[70] [69] La depredación en los pantanos está negativamente relacionada con la profundidad del agua bajo el nido.[70] Las colonias situadas en pantanos son más grandes y tienden a ser más sincrónicas que las ubicadas en tierras altas; sin embargo, la duración de la actividad vinculada a la anidación es mayor en colonias de grandes.[74] En las tierras altas, las dimensiones de las colonias podrían estar restringidas por la escasez de determinados recursos, tales como los sitios de nidificación y el alimento.[74] Los pantanos son generalmente más productivos que los hábitats de tierras altas y presentan una fuente de alimento —insectos acuáticos emergentes— que es renovada rápida y continuamente.[69] El gran tamaño y la nidificación sincrónica de algunas colonias reduce la depredación desde una perspectiva porcentual.[74] Las hembras tienden a construir sus nidos en sitios próximos a puntos de observación desde los que los machos pueden vigilarlos. Los nidos exitosos suelen estar asociados a puntos de observación significativamente más altos y cercanos que los que sufren ataques de depredadores. No existe evidencia, sin embargo, de que los machos que se ocupan más de vigilar sus nidos sean más agresivos en su defensa o que experimenten un mayor éxito respecto de la obtención de parejas o un mayor éxito reproductivo que aquellos que se dedican menos a esa tarea.[28] En vista de la restringida incidencia del parasitismo de puesta del tordo cabecicafé (Molothrus ater) en nidos de tordos sargento y su aparentemente insignificante efecto sobre la mortalidad de los polluelos, parecería que este factor merece poca consideración respecto del éxito reproductivo del tordo sargento.[76]
Según un estudio realizado en las praderas de pasto alto del sur de Wisconsin, la quema controlada tiene escaso impacto sobre la reproducción del tordo sargento. Las quemas de primavera no retrasaron el comienzo de la temporada reproductiva ni afectaron la cantidad de nidos construidos.[77]
Depredación y parasitismo

Probablemente el hombre mate a más tordos sargento que cualquier depredador natural debido a que éstos atacan los cultivos de cereales.[25] Entre los depredadores conocidos de la especie, se encuentran el mapache (Procyon lotor),[22] [43] el visón americano (Neovison vison),[22] la urraca (Pica pica),[22] [43] el chivirín pantanero (Cistothorus palustris),[22] [25] los búhos,[22] los halcones,[22] las serpientes —como la serpiente de agua del norte (Nerodia sipedon)—,[7] [25] [3] el zanate norteño (Quiscalus quiscula),[25] etc. Los depredadores nocturnos como el mapache no son atacados por los tordos sargento adultos.[43] Asimismo, puede alojar ectoparásitos como distintas especies de ftirápteros (por ejemplo, los ischnóceros Philopterus agelaii y Brueelia ornatissima)[38] y ácaros hematófagos (como Ornithonyssus sylviarum, perteneciente a la familia Macronyssidae)[38] y endoparásitos como Haemoproteus quiscalus,[38] Leucocytozoon icteris,[38] Plasmodium vaughani,[38] nematodos,[38] [78] trematodos[78] y cestodos.[78]
Debido a la depredación, especialmente de los huevos y polluelos, el tordo sargento ha desarrollado diversas adaptaciones para proteger sus nidos. Una de ellas consiste en nidificar en grupos, lo cual reduce el peligro puesto que existe un mayor número de padres alertas. Anidar sobre el agua también disminuye las probabilidades de depredación. Los nidos en particular ofrecen una ventaja estratégica puesto que con frecuencia se hallan ocultos entre carrizos ribereños, ubicados normalmente a una altura de uno o dos metros.[22] Los machos a menudo hacen las veces de centinelas, empleando un repertorio de llamados que denotan la proximidad de la amenaza.[79] En especial los machos persiguen a los depredadores con el fin de ahuyentarlos, incluso a animales de dimensiones muy superiores. La coloración apagada de la hembra puede servir para camuflarla, protegiéndola a ella y a su nido cuando está incubando.[22]
Parasitismo de puesta

El tordo sargento es víctima del parasitismo de puesta, en particular por parte del tordo cabecicafé (Molothrus ater), un ictérido cuya reproducción depende de la puesta de sus huevos en los nidos de otras aves.[25] Aparentemente, en el extremo sur de Texas, los nidos de la subespecie A. p. megapotamus son vulnerados con bastante regularidad por el tordo ojirrojo (Molothrus aenus);[80] también existen registros de nidos de megapotamus parasitados por tordos ojirrojos procedentes del estado mexicano de Tamaulipas.[76]
Se trata del único ictérido frecuentemente parasitado por el tordo cabecicafé.[76] Constituye una especie receptiva, es decir, acepta y empolla los huevos extraños; no los expulsa de su nido como otras aves.[75] [80] [81] Por otro lado, tanto machos como hembras son agresivos hacia los tordos cabecicafé adultos.[75] En un experimento, uno o dos de los miembros de cada pareja reaccionaron agresivamente en el 97,1% de los nidos cerca de los cuales se dispuso un espécimen muerto preservado de tordo cabecicafé hembra adulto.[82] También son agresivos hacia los tordos cabecicafé machos: se sabe de casos en los que tordos sargento machos se unieron en la persecución de ejemplares masculinos de esta ave parasítica por varios cientos de metros.[81] Cabe destacar, sin embargo, que esta hostilidad persiste sólo durante la temporada reproductiva: tordos sargento y cabecicafé forman parte de las mismas bandadas y descansan en conjunto pacíficamente en otoño e invierno, lo cual difiere de lo sucedido con algunas aves hospedadoras del Viejo Mundo que muestran conductas agresivas hacia los parásitos de puesta durante todo el año.[76] En algunas ocasiones, cuando un tordo cabecicafé parasita un nido de tordo sargento antes de que éste haya depositado alguno de sus huevos, el huésped abandona su nido.[76] [83] Los polluelos de tordo cabecicafé en nidos con dos o más polluelos de tordo sargento tienen escasas probabilidades de supervivencia debido al mayor tamaño y la agresividad de sus compañeros de nido.[76]
Las tasas de parasitismo de puesta del tordo cabecicafé en nidos de tordos sargento varían enormemente entre distintas regiones.[80] [75] [76] En el este de Estados Unidos y Canadá rara vez son vulnerados,[80] pero en las paderas centrales de ambos países la incidencia del parasitismo en nidos de esta especie es mayor.[80] [84] Hacia comienzos del siglo XX, la subespecie Agelaius phoeniceus fortis fue considerada uno de los huéspedes más comúnmente parasitados por Molothrus ater ater en el valle de Clear Creek (Colorado).[30] De 99 nidos observados en el condado de Washtenaw (Míchigan) entre 1946 y 1949, 5 (el 5,0%) contenían un huevo de tordo cabecicafé, de los cuales uno logró eclosionar pero no abandonar el nido; no se registraron parasitaciones múltiples.[83] En un estudio realizado en 1978 y entre 1986 y 1988 en el condado de Grant (Washington), el 7,7% de los nidos de tordos sargento estaba parasitado.[75] En el sur de Wisconsin, se encontraron tasas de parasitismo de un 15,1% y un 24,3% en la misma pradera en dos años consecutivos (1995 y 1996, respectivamente); no se registraron parasitaciones múltiples el primer año, mientras que en 1996 el 18,5% de los nidos vulnerados contenía más de un huevo extraño.[82] Otras investigaciones realizadas en diferentes puntos de Estados Unidos y Canadá a lo largo de la segunda mitad del siglo XX han arrojado como resultado tasas de parasitismo extremadamente diversas que variaban entre el 0,0% (en un estudio en pantanos de Ontario) y el 54,2% (en canaletas a los costados de carreteras en Nebraska).[75] En el condado de Decatur (Kansas), se halló, a principios del siglo XX, una incidencia parasítica en nidos de tordo sargento de alrededor de un 90%.[76]
Las tres subespecies de tordo cabecicafé (Molothrus ater ater, M. a. artemisiae y M. a. obscurus) están involucradas en parasitaciones a nidos de tordos sargento. Hacia 1963, se reconocían las siguientes nueve razas de tordo sargento como víctimas del parasitismo del tordo cabecicafé: phoeniceus, littoralis, megapotamus, arctolegus, fortis, nevadensis, californicus, neutralis y sonoriensis. Las combinaciones subespecíficas registradas hasta entonces eran las siguientes: M. a. ater parasitaba nidos de phoeniceus, littoralis, megapotamus, arctolegus y fortis; M. a. artemisiae tenía por huéspedes a arctolegus, fortis, nevadensis y californicus; y, por último, M. a. obscurus vulneraba los nidos de megapotamus, californicus, neutralis, littoralis y sonoriensis.[76]
Efectos del momento de actividad del nido
En el susodicho estudio en el condado de Grant (Washington), la tasa de parasitismo de puesta por parte del tordo cabecicafé se incrementó a medida que avanzaba la temporada reproductiva del tordo sargento: los primeros nidos casi nunca eran vulnerados mientras que los tardíos frecuentemente sufrían la intrusión del parásito. Una posible explicación consiste en que los tordos sargento simplemente no constituyen huéspedes predilectos y que los tordos cabecicafé sincronizan su temporada reproductiva de tal manera que se superponga con la de sus huéspedes preferidos, de nidificación tardía. Otra posibilidad podría ser que este fenómeno esté relacionado con el hecho de que cuando la época de reproducción ya está avanzada son menos las hembras presentes en las colonias que puedan participar en la defensa grupal de los nidos activos. La biología de los polluelos de tordo sargento sugiere una tercera hipótesis. La probabilidad de que los polluelos mueran en el nido disminuye dramáticamente después del primer mes de la temporada reproductiva de esta ave hospedadora. Debido a que su descendencia tiene menos probabilidades de morir de inanición en la época de reproducción tardía, podría ser provechoso para las hembras del parásito depositar sus huevos en esos nidos, incluso siendo mayor entonces el peligro de depredación. Puesto que los tordos sargento son un 50% más grandes que los tordos cabecicafé, los polluelos de esta especie podrían tener pocas posibilidades de sobrevivir al competir con los del huésped durante las primeras semanas de la temporada reproductiva, en las que el alimento es limitado.[75]
Por otro lado, en Kent Island (Maryland), en 1958 y 1959, los pocos casos descubiertos de parasitismo de puesta en nidos de tordos sargento correspondían invariablemente a los primeros nidos de la temporada, lo cual aparentemente se debería a que los tordos sargento comenzaban a anidar antes que la mayoría de las especies de paseriformes y a que los tordos cabecicafé empezaban a depositar sus huevos aproximadamente al mismo tiempo que los tordos sargentos; sin embargo, según esta misma fuente, el tordo cabecicafé no parecía perturbar los nidos del tordo sargento ya más avanzada la temporada reproductiva cuando aumentaba la disponibilidad de nidos de otros huéspedes.[76] De modo similar, una investigación en Wisconsin encontró mayores niveles de parasitismo en nidos de tordo sargento durante la primera parte de la temporada reproductiva, en el apogeo de la reproducción del tordo sargento.[85]
Efectos de la ubicación del nido
En el condado de Grant (Washington), el 21,5% de los casos registrados de parasitismo de puesta en nidos de tordo sargento se produjo en nidos inactivos después de que la nidada hubiera sido aniquilada por depredadores o se hubiera independizado. Debido a que varios de estos nidos inactivos recibieron más de un huevo del intruso, considerablemente más de un huevo sobre cinco fue depositado en un nido en el que no tenía posibilidades de eclosionar. Los pantanos con más árboles presentaban un menor porcentaje de huevos de tordo cabecicafé depositados en nidos inactivos de tordos sargento, aunque el número de árboles no afectaba la tasa de parasitismo total en los distintos pantanos. Esto probablemente se deba a que las hembras de tordo cabecicafé podían posarse en o cerca de los pantanos en presencia de árboles y así evaluar la actividad en los nidos, lo que les permitiría escoger mejor cuál parasitar.[75] Por otro lado, en el sur de Wisconsin los nidos de tordo sargento parasitados eran significativamente más proclives a ubicarse cerca de árboles que los no parasitados. Otras variables, como la distancia a puntos de observación que no fueran árboles —como vegetación no leñosa o postes de cercos, los cuales al ser tan frecuentemente usados por los tordos sargento machos probablemente no fueran muy utilizados por los tordos cabecicafé—, la altura y grado de exposición del nido y el grado de agresividad por parte de los tordos sargento adultos, no incidieron significativamente en las tasas de parasitismo en ese estudio.[82]
En las praderas de pasto alto del sur de Wisconsin, se descubrió que las probabilidades de parasitismo en nidos de tordo sargento aumentaban con la proximidad al borde de un hábitat —en este caso, las áreas entre las praderas y terrenos agrícolas o segmentos de bosque— o a una carretera y con la distancia desde superficies quemadas.[77]
Efectos de la defensa grupal y la nidificación colectiva
Las colonias densas de tordos sargento presentan tasas más bajas de parasitismo, lo que sugiere que la defensa grupal contra los tordos cabecicafé juega un papel importante.[75] [80] [76] Tal como podría esperarse, son justamente las aves que anidan en densidades altas en colonias de pantanos las que participan más a menudo en ataques colectivos hacia el intruso.[81] En regiones en las que los tordos sargento anidan mayormente en colonias en pantanos de totoras (el este de Estados Unidos y Canadá), no constituyen un huésped habitual del tordo cabecicafé; sin embargo, en otras áreas en las que construyen sus nidos de manera más esparcida sobre arbustos, estos son parasitados a menudo.[76] Pareciera que, aunque no puedan penetrar en colonias densas, los tordos cabecicafé son relativamente libres de merodear en hábitats de tierras altas en donde los tordos sargento se encuentran demasiado dispersos para permitir un ataque en conjunto efectivo.[81] Ninguno de los 363 nidos en cinco colonias en pantanos en el área de Kingston (Ontario) en 1972 y 1973 fue parasitado, mientras que el 22,8% de 35 nidos en tierras altas lo fue.[81] Las aves que nidan en densidades altas se benefician de una detección más efectiva de los tordos cabecicafé y de los depredadores, además de las ventajas que proporciona la defensa grupal para ahuyentarlos.[81] Las respuestas de defensa grupal parecen no estar relacionadas a la etapa de nidificación de los individuos involucrados en la persecución, ya que los tordos cabecicafé son atacados colectivamente tanto por machos en cuyos nidos los huevos aún no han eclosionado como por otros que ya tienen polluelos.[81]
En Wisconsin, se examinaron los efectos de la nidificación en colonias de tordos sargento sobre el parasitismo del tordo cabecicafé y se registró una relación positiva significativa entre la cantidad de nidos comenzados por día y la proporción de nidos parasitados, lo que sugería que la nidificación colectiva no repercutía en una menor amenaza de parasitación. No obstante, la sincronización en la puesta de huevos aparentemente contribuía a reducir el riesgo de parasitismo. Los nidos vulnerados tenían períodos de puesta menos sincronizados con los de sus vecinos más cercanos y se encontraban más alejados del nido simultáneamente activo más próximo —es decir, estaban más aislados— que en el caso de los nidos sin parasitar.[85] En un estudio en Míchigan en el que se encontró una tasa parasítica de 1 nido sobre 185, se descubrió que todos los nidos parasitados estaban aislados o en los perímetros de las colonias, a treinta metros o más del nido de tordo sargento más cercano. Esto se debe a que es posible que un tordo cabecicafé aove en un nido solitario sin ser descubierto y perseguido, no así en una colonia.[76]
Una manera de determinar si la baja incidencia del parasitismo sobre esta especie en algunas áreas se debe exclusivamente a la defensa grupal de los nidos o si una escasa predilección por el tordo sargento como huésped de los huevos del intruso constituye también un factor a tener en cuenta consiste en evaluar el grado de parasitismo sufrido por nidos de otras especies que aniden entre las colonias de tordos sargento. Si la defensa del nido es la única explicación para la baja incidencia parasítica que disfrutaban los tordos sargento, las demás especies que se reprodujeran junto a esta deberían también experimentar niveles reducidos de parasitismo pues la "defensa comunal" debería mantener a raya a los tordos cabecicafé respecto de toda el área. Sin embargo, se tiene noticia de casos en los que chipes de cara negra (Geothlypis trichas) y gorriones coronirrufos pantaneros (Melospiza georgiana) sufrían niveles altos de parasitismo en sitios en los que los nidos de tordos sargento no habían sido vulnerados en absoluto o lo habían sido en una baja proporción. Esto apoya la hipótesis de que las preferencias del parásito respecto de otras aves hospedadoras influyen sobre la escasa cantidad de nidos de tordos sargento parasitados en ciertas regiones.[80] No obstante ello, otras observaciones sugieren que la tendencia de los tordos sargento a ahuyentar a estos intrusos efectivamente repercute en una menor tasa parasítica en nidos cercanos de víreos y chipes.[76]
Impacto ecológico y económico
 Macho posado en una espiga.
Macho posado en una espiga.Los tordos sargento pueden tener un impacto importante y duradero sobre el medio ambiente. A través del control de las poblaciones de insectos por medio de la predación y de las hierbas indeseadas por medio del consumo de sus semillas, permiten el crecimiento de plantas y cultivos de mayores dimensiones.[22] Se alimentan de Anthonomus grandis e Hypera postica, dos especies de curculiónidos que afectan el algodón y la alfalfa respectivamente.[25]
Paradójicamente, también pueden devastar los campos de cultivo. A pesar de que consumen semillas de malezas, son conocidos por ocasionar grandes daños a la agricultura a causa de sus hábitos de descansar en grupos masivos y su gusto por los productos agrícolas. Pueden ser perjudiciales para los campos de maíz, arroz y girasol.[22] De hecho, en América del norte, el daño a los cultivos de maíz por parte de esta especie aumentó desde finales de la década de 1960 a principios de la de 1980, quizá debido al incremento del área destinada a la producción de granos[29] [37] [86] y por la disminución de superficie ocupada por pequeñas áreas con rastrojo, henares y terrenos no cultivados, lo cual, a su vez, aumentó la dependencia del ave con respecto al maíz para asegurarse el alimento.[86] [87] De 215 especies neotropicales migratorias, Rodenhouse y otros (1992) lo identificaron como la que producía, por un amplio margen, la mayor pérdida económica.[86]
Esto ha llevado a la utilización de trampas, veneno y tensoactivos por parte de los agricultores en un intento por controlar las poblaciones de tordos sargento. Estos últimos eliminan las propiedades impermeables de las plumas, haciéndolos extremadamente vulnerables a las bajas temperaturas.[22] Métodos menos nocivos incluyen la eliminación de los residuos de las cosechas, los cuales atraen las aves a los campos.[22]
 Bandada volando en el crepúsculo.
Bandada volando en el crepúsculo.En las regiones septentrionales de su área de distribución no representan un problema a gran escala, pues allí su dieta se centra en las semillas de hierbas e insectos. No obstante, provocan el detrimento de los granos que germinan en primavera y del maíz dulce en verano rasgando la cobertura foliácea de las mazorcas y arruinándolas desde un punto de vista comercial. También atacan otros granos de manera limitada, pero la mayor parte de los que consumen son desperdicios que quedaron en el suelo. En el Medio Oeste, donde estas aves son mucho más abundantes que en el norte y donde hay extensos cultivos de cereales, los tordos sargento y otros ictéridos, a fines del verano y en otoño, causan gran daño a los campos de granos, tanto mientras éstos están madurando como cuando son cosechados. Sin embargo, se ha afirmado que incluso allí resultan beneficiosos debido a las larvas perniciosas que eliminan de las espigas, pudiendo contrarrestar plagas de orugas. En los estados sureños, perjudican gravemente al arroz arrancando los plantines en primavera y comiendo los granos mientras maduran, siendo en este sentido casi tan dañinos como los tordos arroceros (Dolichonyx oryzivorus). Por otro lado, son de cierta utilidad al consumir las semillas de malezas que de otro modo devaluarían el producto.[25]
 Macho posado en un tronco.
Macho posado en un tronco.Beal (1900) afirmó: “Al resumir el estatus económico del tordo sargento, el principal aspecto a tener en cuenta es el pequeño porcentaje que representan los granos en su dieta anual, lo cual aparentemente contradice las quejas respecto de sus hábitos destructivos. A juzgar por los contenidos estomacales, el tordo sargento es decididamente un ave útil. El servicio que brinda a través de la eliminación de insectos y semillas de hierbas nocivos supera ampliamente los daños que produce su consumo de granos. Los estragos que a veces provoca deberían ser atribuidos enteramente a su excesiva abundancia en ciertos lugares.”[25] Por otro lado, Bendell y otros (1981) encontraron que el beneficio económico del control de plagas, tales como el taladro del maíz (Ostrinia nubilalis), por parte del tordo sargento sólo compensaba el 20% del daño a los cultivos ocasionados por esta ave.[86] [88]
Al ser una de las aves más numerosas del continente, juega un rol importante en la dispersión de otras especies. Puesto que los tordos se agrupan y descansan en cantidades tan grandes, la supervivencia de determinadas especies que se unen a sus bandadas seguramente se vea afectada por su compañía. En síntesis, estas aves son tan numerosas y activas que su sola presencia y comportamiento natural es suficiente para influir sobre el medio ambiente de un modo visible.[22]
Referencias
- ↑ a b c BirdLife International (2008). «Agelaius phoeniceus». Lista Roja de especies amenazadas de la UICN 2010.4.. Consultada: 22 de diciembre 2010.
- ↑ a b c d «ITIS Standard Report Page: Agelaius phoeniceus» (en inglés). Integrated Taxonomic Information System. Consultado el 22 de diciembre de 2010.
- ↑ a b c d e «Agelaius phoeniceus phoeniceus» (en español). Universidad Francisco Marroquín. Consultado el 10 de enero de 2011.
- ↑ a b c d e f g h i j k l m n ñ o p q r s t u v w «Red-winged Blackbird» (en inglés). All About Birds. Ithaca (Nueva York): Cornell Lab of Ornithology (2009). Consultado el 23 de diciembre de 2010.
- ↑ a b c d e f g h i Bologna, Gianfranco (1981) (en español). Guías de la naturaleza: Aves. Barcelona: Grijalbo. ISBN 84-253-3362-8.
- ↑ a b c d e f g h Orians, Gordon H. (1980) (en inglés). Some adaptations of marsh-nesting blackbirds. Princeton (Nueva Jersey): Princeton University Press. ISBN 0691082375. http://books.google.com.ar/books?hl=es&lr=&id=Of4lIyjpBF0C&oi=fnd&pg=PA3&dq=Some+adaptations+of+marsh-nesting+blackbirds&ots=3bkSuBEIEE&sig=eVG-yUMALrH7rHs6wrd-TwImRgc#v=onepage&q&f=false. Consultado el 12 de agosto de 2011.
- ↑ a b c d e f g h i j k l m n ñ o p q r s t u v w x y z «Red-winged Blackbirds: Species information and Photos» (en inglés). Consultado el 24 de diciembre de 2010.
- ↑ «Catalogue of Life: 2010 Annual Checklist» (en inglés). Integrated Taxonomic Information System. Consultado el 22 de enero de 2011.
- ↑ a b Lowther, P. E.; Fraga, R.; Schulenberg, T. S.; Lanyon, S. M. (2004). «Nomenclatural solution for a polyphyletic Agelaius» (en inglés). Bulletin of the British Ornithologists' Club (British Ornithologists' Club) 124 (3): p. 171-177. http://grupofalco.com.ar/pedefes/Lowther_Agelasticus.pdf. Consultado el 20 de julio de 2011.
- ↑ Vieillot, L.P. (1816) Analyse D'Une Nouvelle Ornithologie Elementaire p. 33
- ↑ a b c «Red-Winged Blackbird (Agelaius phoeniceus)» (en inglés). National Geographic Complete Birds of North America. National Geographic Society (2006). Consultado el 20 de julio de 2011.
- ↑ a b Whittingham, Linda A.; Kirkconnell, Arturo; Ratcliffe, Laurene M. (octubre de 1992). «Differences in song and sexual dimorphism between Cuban and North American red-winged blackbirds (Agelaius phoeniceus)» (en inglés). The Auk (American Ornithologists' Union) 109 (4): p. 928-933. http://www.jstor.org/pss/4088178. Consultado el 20 de julio de 2011.
- ↑ a b Garrido, Orlando; Kirkconnell, Arturo (junio de 1996). «Taxonomic status of the Cuban form of the red-winged blackbird» (en inglés). The Wilson Bulletin (Wilson Ornithological Society) 108 (2): p. 372-374. http://www.jstor.org/pss/4163690. Consultado el 19 de julio de 2011.
- ↑ Whittingham, Linda A.; Kirkconnell, Arturo; Ratcliffe, Laurene M. (noviembre de 1996). «Breeding behavior, social organization and morphology of red-shouldered (Agelaius assimilis) and tawny-shouldered (Agelaius humeralis) blackbirds» (en inglés). The Condor (Cooper Ornithological Society) 98 (4): p. 832-836. http://elibrary.unm.edu/sora/Condor/files/issues/v098n04/p0832-p0836.pdf. Consultado el 10 de julio de 2011.
- ↑ Searcy, William A. (1979). «Morphological correlates of dominance in captive male red-winged blackbirds» (en inglés). The Condor (Cooper Ornithological Society) 81: p. 417-420. http://elibrary.unm.edu/sora/Condor/files/issues/v081n04/p0417-p0420.pdf. Consultado el 30 de agosto de 2011.
- ↑ a b c Johnson, Kevin P.; Lanyon, Scott M. (2000). «Evolutionary changes in color patches of blackbirds are associated with marsh nesting» (en inglés). Behavioral Ecology (International Society for Behavioural Ecology) 11 (5): p. 515-519. http://beheco.oxfordjournals.org/content/11/5/515.full.pdf+html. Consultado el 11 de agosto de 2011.
- ↑ a b c d Shutler, Dave; Weatherhead, Patrick J. (1991). «Owner and floater red-winged blackbirds: determinants of status» (en inglés). Behavioral Ecology and Sociobiology (Springer-Verlag) 28 (4): p. 235-241. http://www.acadiau.ca/~dshutler/1991BES.pdf. Consultado el 20 de agosto de 2011.
- ↑ Røskaft, Eivin; Rohwer, Sievert (agosto de 1987). «An experimental study of the function of the red epaulettes and the black body colour of male red-winged blackbirds» (en inglés). Animal Behaviour (Association for the Study of Animal Behaviour) 35 (4): p. 1070-1077. http://www.sciencedirect.com/science/article/pii/S0003347287801641. Consultado el 2 de septiembre de 2011.
- ↑ Smith, Douglas G. (1972). «The role of the epaulets in the red-winged blackbird, (Agelaius phoeniceus) social system» (en inglés). Behaviour (BRILL) 41 (3/4): p. 251-268. http://www.jstor.org/stable/4533440. Consultado el 20 de agosto de 2011.
- ↑ Crawford, Richard D. (marzo de 1977). «Breeding biology of year-old and older female red-winged and yellow-headed blackbirds» (en inglés). The Wilson Bulletin (Wilson Ornithological Society) 89 (1): p. 73-80. http://www.jstor.org/stable/4160871. Consultado el 26 de septiembre de 2011.
- ↑ a b c Gough, Gregory (2003). «Red-winged blackbird Agelaius phoeniceus» (en inglés). USGS Patuxent Wildlife Research Center. Consultado el 23 de diciembre de 2010.
- ↑ a b c d e f g h i j k l m n ñ o p q r s t u v w x y z aa ab ac ad ae Rosenthal, Anthony (2004). «ADW: Agelaius phoeniceus: Information» (en inglés). Animal Diversity Web. University of Michigan Museum of Zoology. Consultado el 24 de diciembre de 2010.
- ↑ Irwin, Rebecca E. (diciembre de 1994). «The evolution of plumage dichromatism in the New World Blackbirds: social selection on female brightness» (en inglés). The American Naturalist (American Society of Naturalists) 144 (6): p. 890-907. http://www.jstor.org/stable/2463134. Consultado el 13 de agosto.
- ↑ a b c Searcy, William A. (junio de 1979). «Sexual selection and body size in male red-winged blackbirds» (en inglés). Evolution (Society for the Study of Evolution) 33 (2): p. 649-661. http://www.jstor.org/stable/2407788. Consultado el 17 de agosto de 2011.
- ↑ a b c d e f g h i j k l m n ñ o p q r s t u v w x y z aa Bent, Arthur Cleveland (1958). «Red-winged Blackbird, Agelaius phoeniceus [Eastern Redwing]» (en inglés). Life Histories of North American Blackbirds, Orioles, Tanagers, and Allies (Life Histories of North American Birds). Instituto Smithsoniano, United States National Museum Bulletin 211. pp. 123-150. http://birdsbybent.netfirms.com/ch41-50/blackbir.html. Consultado el 20 de febrero de 2011.
- ↑ Clark, W. Dwight; Karr, James R. (marzo de 1979). «Effects of highways on red-winged blackbird and horned lark populations» (en inglés). The Wilson Bulletin (Wilson Ornithological Society) 91 (1): p. 143-145. http://elibrary.unm.edu/sora/Wilson/v091n01/p0143-p0145.pdf. Consultado el 5 de septiembre.
- ↑ Weatherhead, Patrick J.; Hoysak, Drew J. (julio de 1984). «Dominance structuring of a red-winged blackbird roost» (en inglés). The Auk (American Ornithologists' Union) 101 (3): p. 551-555. http://www.jstor.org/stable/4086608. Consultado el 4 de septiembre de 2011.
- ↑ a b Yasukawa, Ken; Whittenberger, Lynn K.; Nielsen, Tracy A. (junio de 1992). «Anti-predator vigilance in the red-winged blackbird, Agelaius phoeniceus: do males act as sentinels?» (en inglés). Animal Behaviour (Association for the Study of Animal Behaviour) 43 (6): p. 961-969. http://www.sciencedirect.com/science/article/pii/S0003347206800096. Consultado el 10 de octubre de 2011.
- ↑ a b c White, Stephen B.; Dolbeer, Richard A.; Bookhout, Theodore A. (abril de 1985). «Ecology, bioenergetics, and agricultural impacts of a winter-roosting population of blackbirds and starlings» (en inglés). Wildlife Monographs (Allen Press) (93): p. 3-42. ISSN 0084-0173. http://www.jstor.org/stable/3830662. Consultado el 9 de agosto de 2011.
- ↑ a b Lincoln, Frederick Charles (enero de 1920). «Birds of the Clear Creek district, Colorado» (en inglés). The Auk (American Ornithologists' Union) 37 (1): p. 60-77. http://elibrary.unm.edu/sora/Auk/v037n01/p0060-p0077.pdf. Consultado el 10 de octubre de 2011.
- ↑ a b c d e f g Shutler, Dave; Weatherhead, Patrick J. (1991). «Basal song rate variation in male red-winged blackbirds: sound and fury signifying nothing?» (en inglés). Behavioral Ecology (International Society for Behavioural Ecology) 2 (2): p. 123-132. http://www.acadiau.ca/~dshutler/1991BehavEcol.pdf. Consultado el 8 de septiembre de 2011.
- ↑ Yasukawa, Ken; Blank, J. L.; Patterson, C. B. (1980). «Song repertoires and sexual selection in the red-winged blackbird» (en inglés). Behavioral Ecology and Sociobiology (Springer-Verlag) 7 (3): p. 233-238. http://www.springerlink.com/content/q0648t127u246840/. Consultado el 23 de septiembre de 2011.
- ↑ a b c d Searcy, William A.; Yasukawa, Ken (1990). «Use of the song repertoire in intersexual and intrasexual contexts by male red-winged blackbirds» (en inglés). Behavioral Ecology and Sociobiology (Springer-Verlag) 27 (2): p. 123-128. http://www.jstor.org/stable/4600454. Consultado el 9 de agosto de 2011.
- ↑ a b Yasukawa, Ken (febrero de 1981). «Song repertoires in the red-winged blackbird (Agelaius phoeniceus): A test of the Beau Geste hypothesis» (en inglés). Animal Behaviour (Association for the Study of Animal Behaviour) 29 (1): p. 114-125. http://www.sciencedirect.com/science/article/pii/S0003347281801583. Consultado el 23 de septiembre de 2011.
- ↑ Yasukwa, Ken (noviembre de 1989). «The costs and benefits of a vocal signal: the nest-associated ‘Chit’ of the female red-winged blackbird, Agelaius phoeniceus» (en inglés). Animal Behaviour (Association for the Study of Animal Behaviour) 38 (5): p. 866-874. http://www.sciencedirect.com/science/article/pii/S0003347289801186. Consultado el 10 de octubre de 2011.
- ↑ Yasukawa, Ken; Boley, Rebecca A.; Simon, Sue E. (octubre de 1987). «Seasonal change in the vocal behaviour of female red-winged blackbirds, Agelaius phoeniceus» (en inglés). Animal Behaviour (Association for the Study of Animal Behaviour) 35 (5): p. 1416-1423. http://www.sciencedirect.com/science/article/pii/S0003347287800143. Consultado el 10 de octubre de 2011.
- ↑ a b Clark, Robert G.; Weatherhead, Patrick J.; Greenwood, Hamilton; Titman, Rodger D. (1986). «Numerical responses of red-winged blackbird populations to changes in regional land-use patterns» (en inglés). Canadian Journal of Zoology (NRC Research Press) 64 (9): p. 1944-1950. http://www.nrcresearchpress.com/doi/abs/10.1139/z86-292. Consultado el 9 de agosto de 2011.
- ↑ a b c d e f g h Dufour, Kevin W.; Weatherhead, Patrick J. (1998). «Bilateral symmetry as an indicator of male quality in red-winged blackbirds: associations with measures of health, viability, and parental effort» (en inglés). Behavioral Ecology (International Society for Behavioural Ecology) 9 (3): p. 220-231. http://beheco.oxfordjournals.org/content/9/3/220.full.pdf+html. Consultado el 10 de agosto de 2011.
- ↑ a b c Weatherhead, Patrick J.; Dufour, Kevin W.; Lougheed, Stephen C.; Eckert, Christopher G. (1999). «A test of the good-genes-as-heterozygosity hypothesis using red-winged blackbirds» (en inglés). Behavioral Ecology (International Society for Behavioural Ecology) 10 (6): p. 619-625. http://beheco.oxfordjournals.org/content/10/6/619.full. Consultado el 13 de agosto de 2011.
- ↑ a b c d e f g h i j k l m n Dufour, Kevin W.; Weatherhead, Patrick J. (1998). «Reproductive consequences of bilateral asymmetry for individual male red-winged blackbirds» (en inglés). Behavioral Ecology (International Society for Behavioural Ecology) 9 (3): p. 232-242. http://beheco.oxfordjournals.org/content/9/3/232.full.pdf+html. Consultado el 15 de agosto de 2011.
- ↑ Dufour, Kevin W.; Weatherhead, Patrick J. (1998). «Bilateral symmetry and social dominance in captive male red-winged blackbirds» (en inglés). Behavioral Ecology and Sociobiology (Springer-Verlag) 42 (1): p. 71-76. http://www.springerlink.com/content/3xrx0h45u0rk8ta2/. Consultado el 11 de agosto de 2011.
- ↑ Weatherhead, Patrick J. (1990). «Secondary sexual traits, parasites, and polygyny in red-winged blackbirds, Agelaius phoeniceus» (en inglés). Behavioral Ecology (International Society for Behavioural Ecology) 1 (2): p. 125-130. http://beheco.oxfordjournals.org/content/1/2/125.abstract. Consultado el 22 de septiembre de 2011.
- ↑ a b c d e f g h Weatherhead, Patrick J. (1995). «Effects on female reproductive success of familiarity and experience among male red-winged blackbirds» (en inglés). Animal Behaviour (Association for the Study of Animal Behaviour) 49: p. 967–976. http://www.life.illinois.edu/pjw/reprints/1995-W-AB.pdf. Consultado el 17 de agosto de 2011.
- ↑ a b Yasukawa, Ken (agosto de 1979). «Territory establishment in red-winged blackbirds: importance of aggressive behavior and experience» (en inglés). The Condor (Cooper Ornithological Society) 81 (3): p. 258-264. http://www.jstor.org/stable/1367628. Consultado el 19 de agosto de 2011.
- ↑ a b Searcy, William A.; Yasukawa, Ken; Lanyon, Scott (enero de 1999). «Evolution of polygyny in the ancestors of red-winged blackbirds» (en inglés). The Auk (American Ornithologists' Union) 116 (1): p. 5-19. http://www.jstor.org/stable/4089449. Consultado el 10 de agosto de 2011.
- ↑ Wiley, R. Haven; Hartnett, Sue A. (mayo de 1976). «Effects of interactions with older males on behavior and reproductive development in first-year male red-winged blackbirds Agelaius phoeniceus». Journal of Experimental Zoology (Wiley-Blackwell) 196 (2): p. 231-242. http://onlinelibrary.wiley.com/doi/10.1002/jez.1401960210/abstract. Consultado el 31 de agosto de 2011.
- ↑ a b c Holcomb, Larry C.; Twiest, Gilbert (septiembre de 1970). «Growth rates and sex ratios of red-winged blackbird nestlings» (en inglés). The Wilson Bulletin (Wilson Ornithological Society) 82 (3): p. 294-303. http://www.jstor.org/stable/4159981. Consultado el 30 de septiembre de 2011.
- ↑ Wright, Philip L.; Wright, Margaret H. (marzo-abril de 1944). «The reproductive cycle of the male red-winged blackbird» (en inglés). The Condor (Cooper Ornithological Society) 46 (2): p. 46-59. http://www.jstor.org/stable/1364305. Consultado el 13 de septiembre de 2011.
- ↑ a b c d e f g Shutler, Dave; Weatherhead, Parick J. (1994). «Movement patterns and territory acquisition by male red-winged blackbirds» (en inglés). Canadian Journal of Zoology (NRC Research Press) 72: p. 712-720. http://www.acadiau.ca/~dshutler/1994RWBB.pdf. Consultado el 4 de septiembre de 2011.
- ↑ a b c Nero, Robert W. (junio de 1956). «A behavior study of the red-winged blackbird: II. Territoriality» (en inglés). The Wilson Bulletin (Wilson Ornithological Society) 68 (2): p. 129-150. http://www.jstor.org/stable/4158485. Consultado el 7 de septiembre de 2011.
- ↑ Picman, Jaroslav (julio de 1987). «Territory establishment, size, and tenacity by male red-winged blackbirds» (en inglés). The Auk (American Ornithologists' Union) 104 (3): p. 405-412. http://www.jstor.org/stable/4087537. Consultado el 7 de septiembre de 2011.
- ↑ Weatherhead, Patrick J.; Montgomerie, Robert; Gibbs, H. Lisle; Boag, Peter T. (22 de diciembre de 1994). «The cost of extra-pair fertilizations to female red-winged blackbirds» (en inglés). Proceedings: Biological Sciences (The Royal Society) 258 (1353): p. 315-320. http://www.jstor.org/stable/50097. Consultado el 26 de agosto de 2011.
- ↑ Metz, Karen J.; Weatherhead, Patrick J. (1991). «Color bands function as secondary sexual traits in male red-winged blackbirds» (en inglés). Behavioral Ecology and Sociobiology (Springer-Verlag) 28 (1): p. 23-27. http://www.springerlink.com/content/yg7260142011429w/. Consultado el 19 de septiembre de 2011.
- ↑ Peek, Frank W. (diciembre de 1971). «Seasonal change in the breeding behavior of the male red-winged blackbird» (en inglés). The Wilson Bulletin (Wilson Ornithological Society) 83 (4): p. 383-395. http://www.jstor.org/stable/4160135. Consultado el 20 de septiembre de 2011.
- ↑ a b Picman, Jaroslav (diciembre de 1981). «The adaptive value of polygyny in marsh-nesting red-winged blackbirds; renesting, territory tenacity, and mate fidelity of females» (en inglés). Canadian Journal of Zoology (NRC Research Press) 59 (12): p. 2284-2296. http://www.nrcresearchpress.com/doi/abs/10.1139/z81-307. Consultado el 12 de agosto de 2011.
- ↑ Beletsky, Les D.; Orians, Gordon H. (1987). «Territoriality among male red-winged blackbirds: I. Site fidelity and movement patterns» (en inglés). Behavioral Ecology and Sociobiology (Springer-Verlag) 20 (1): p. 21-34. http://www.springerlink.com/content/wq87nr3747841121/. Consultado el 11 de agosto de 2011.
- ↑ a b Yasukawa, Ken; Searcy, William A. (octubre de 1981). «Nesting synchrony and dispersion in red-winged blackbirds: is the harem competitive or cooperative?» (en inglés). The Auk (American Ornithologists' Union) 98 (4): p. 659-668. http://www.jstor.org/stable/4085889. Consultado el 9 de agosto de 2011.
- ↑ Eckert, Christopher G.; Weatherhead, Patrick J. (1987). «Competition for territories in red-winged blackbirds: Is resource-holding potential realized?» (en inglés). Behavioral Ecology and Sociobiology (Springer-Verlag) 20 (5): p. 369-375. http://www.springerlink.com/content/t483r84727111p43/. Consultado el 29 de agosto de 2011.
- ↑ a b Weatherhead, Patrick J.; Boag, Peter T. (1995). «Pair and extra-pair mating success relative to male quality in red-winged blackbirds» (en inglés). Behavioral Ecology and Sociobiology (Springer-Verlag) 37 (2): p. 81-91. http://www.springerlink.com/content/p0vg10t6jh1q104r/. Consultado el 10 de agosto de 2011.
- ↑ a b Yasukawa, Ken; Knight, Richard L.; Knight Skagen, Susan (octubre de 1987). «Is courtship intensity a signal of male parental care in red-winged blackbirds (Agelaius phoeniceus)?» (en inglés). The Auk (American Ornithologists' Union) 104 (4): p. 628-634. http://www.jstor.org/stable/4087273. Consultado el 24 de septiembre de 2011.
- ↑ Searcy, William A. (abril de 1979). «Male characteristics and pairing success in red-winged blackbirds» (en inglés). The Auk (American Ornithologists' Union) 96 (2): p. 353-363. http://www.jstor.org/stable/4085314. Consultado el 17 de agosto de 2011.
- ↑ a b c Gibbs, H. Lisle; Weatherhead, Patrick J.; Boag, Peter T.; White, Bradley N.; Tabak, Lisa M.; Hoysak, Drew J. (7 de diciembre de 1990). «Realized reproductive success of polygynous red-winged blackbirds revealed by DNA markers» (en inglés). Science (Asociación Estadounidense para el Avance de la Ciencia) 250 (4986): p. 1394-1397. http://www.sciencemag.org/content/250/4986/1394.short. Consultado el 16 de agosto de 2011.
- ↑ Westneat, David F. (1993). «Polygyny and extrapair fertilizations in eastern red-winged blackbirds (Agelaius phoeniceus)» (en inglés). Behavioral Ecology (International Society for Behavioural Ecology) 4 (1): p. 49-60. http://beheco.oxfordjournals.org/content/4/1/49.short. Consultado el 19 de agosto de 2011.
- ↑ Weatherhead, Patrick J.; Boag, Peter T. (abril de 1997). «Genetic estimates of annual and lifetime reproductive success in male red-winged blackbirds» (en inglés). Ecology (Ecological Society of America) 78 (3): p. 884-896. http://www.esajournals.org/doi/abs/10.1890/0012-9658%281997%29078%5B0884%3AGEOAAL%5D2.0.CO%3B2?journalCode=ecol. Consultado el 19 de agosto de 2011.
- ↑ Gray, Elizabeth M. (1996). «Female control of offspring paternity in a western population of red-winged blackbirds (Agelaius phoeniceus)» (en inglés). Behavioral Ecology and Sociobiology (Springer-Verlag) 38 (4): p. 267-278. http://www.springerlink.com/content/q23a50efdlvjj1ku/. Consultado el 16 de agosto de 2011.
- ↑ Hergenrader, Gary L. (enero de 1962). «The incidence of nest parasitism by the brown-headed cowbird (Molothrus ater) on roadside nesting birds in Nebraska» (en inglés). The Auk (American Ornithologists' Union) 79 (1): p. 85-88. http://www.jstor.org/stable/4082452. Consultado el 6 de octubre de 2011.
- ↑ a b c d Fiala, Kent L. (1981). «Sex ratio constancy in the red-winged blackbird» (en inglés). Evolution (Society for the Study of Evolution) 35 (5): p. 898-910. http://www.faintlake.com/pubs/SexRatioConstancy.pdf. Consultado el 6 de septiembre de 2011.
- ↑ a b c d e f g h i Fiala, Kent L. (1981). «Reproductive cost and the sex ratio in red-winged blackbirds». En R. D. Alexander y D. W. Tinkle (en inglés). Natural Selection and Social Behavior: Recent Research and New Theory. Chiron Press. http://www.faintlake.com/pubs/ReproductiveCostAndTheSexRatioInRed-wingedBlackbirds.pdf. Consultado el 27 de septiembre de 2011.
- ↑ a b c d e f Robertson, Raleigh J. (junio de 1973). «Optimal niche space of the red-winged blackbird. III. Growth rate and food of nestlings in marsh and upland habitat» (en inglés). The Wilson Bulletin (Wilson Ornithological Society) 85 (2): p. 209-222. http://www.jstor.org/stable/4160323. Consultado el 3 de octubre de 2011.
- ↑ a b c Robertson, Raleigh J. (1972). «Optimal niche space of the redwinged blackbird (Agelaius phoeniceus). I. Nesting success in marsh and upland habitat» (en inglés). Canadian Journal of Zoology (NRC Research Press) 50 (2): p. 247-263. http://www.nrcresearchpress.com/doi/abs/10.1139/z72-036. Consultado el 30 de septiembre de 2011.
- ↑ a b c d e Olson, John M. (1994). «The ontogeny of shivering thermogenesis in the red-winged blackbird (Agelaius phoeniceus)» (en inglés). The Journal of Experimental Biology (Cambridge (Reino Unido): The Company of Biologists) 191: p. 59-88. http://jeb.biologists.org/content/191/1/59.full.pdf+html. Consultado el 10 de agosto de 2011.
- ↑ a b c d Olson, John M. (2001). «Ontogeny of catabolic and morphological properties of skeletal muscle of the red-winged blackbird (Agelaius phoeniceus)» (en inglés). Journal of Comparative Physiology B (Springer-Verlag) 171: p. 527-542. http://www.springerlink.com/content/dqbcbamcqnclgrcy/. Consultado el 10 de agosto de 2011.
- ↑ a b Hill, Richard W.; Beaver, Donald L. (julio de 1982). «Inertial thermostability and thermoregulation in broods of redwing blackbirds» (en inglés). Physiological Zoology (University of Chicago Press) 55 (3): p. 250-266. http://www.jstor.org/stable/30157889. Consultado el 3 de octubre de 2011.
- ↑ a b c d Robertson, Raleigh J. (septiembre de 1973). «Optimal niche space of the redwinged blackbird: Spatial and temporal patterns of nesting activity and success» (en inglés). Ecology (Ecological Society of America) 54 (5): p. 1085-1093. http://www.jstor.org/stable/1935574. Consultado el 1 de octubre de 2011.
- ↑ a b c d e f g h i Freeman, Scott; Gori, David F.; Rohwer, Sievert (1990). «Red-winged blackbirds and brown-headed cowbirds: Some aspects of a host-parasite relationship» (en inglés). The Condor (Cooper Ornithological Society) 92: p. 336-340. http://elibrary.unm.edu/sora/Condor/files/issues/v092n02/p0336-p0340.pdf. Consultado el 8 de agosto de 2011.
- ↑ a b c d e f g h i j k l m n Friedmann, Herbert (1963) (en inglés). Host Relations of the Parasitic Cowbirds. 233. Bulletin of the United States National Museum. p. 1-276. http://hdl.handle.net/10088/10099. Consultado el 6 de octubre de 2011.
- ↑ a b Clotfelter, Ethan D.; Yasukawa, Ken; Newsome, Richard D. (1999). «The effects of prescribed burning and habitat edges on brown-headed cowbird parasitism of red-winged blackbirds» (en inglés). Studies in Avian Biology 18: p. 275-281. http://www3.amherst.edu/~edclotfelter/research_page/research_pdfs/Clotfelter_et_al._SAB_1999.pdf. Consultado el 9 de agosto de 2011.
- ↑ a b c Weatherhead, Patrick J.; Metz, Karen J.; Bennett, Gordon F.; Irwin, Rebecca E. (1993). «Parasite faunas, testosterone and secondary sexual traits in male red-winged blackbirds» (en inglés). Behavioral Ecology and Sociobiology (Springer-Verlag) 33 (1): p. 13-23. http://www.springerlink.com/content/x78gj16568184185/. Consultado el 4 de septiembre de 2011.
- ↑ Beletsky, Les D. (agosto de 1991). «Alert calls of male red-winged blackbirds: call rate and function» (en inglés). Canadian Journal of Zoology (NRC Research Press) 69 (8): p. 2116-2120. http://www.nrcresearchpress.com/doi/abs/10.1139/z91-295. Consultado el 20 de agosto de 2011.
- ↑ a b c d e f g Friedmann, Herbert; Kiff, Lloyd F.; Rothstein, Stephen I. (1977). «A further contribution to knowledge of the host relations of the parasitic cowbirds» (en inglés). Smithsonian Contributions to Zoology (Washington D.C.: Smithsonian Institution Press) 235: p. 5-10. http://www.sil.si.edu/smithsoniancontributions/Zoology/pdf_hi/SCTZ-0235.pdf. Consultado el 26 de junio de 2011.
- ↑ a b c d e f g Robertson, Raleigh J.; Norman, Richard F. (1976). «Behavioral defenses to brood parasitism by potential hosts of the brown-headed cowbird» (en inglés). The Condor (Cooper Ornithological Society) 78 (2): p. 166-173. http://elibrary.unm.edu/sora/Condor/files/issues/v078n02/p0166-p0173.pdf. Consultado el 8 de octubre de 2011.
- ↑ a b c Clotfelter, Ethan D. (1998). «What cues do brown-headed cowbirds use to locate red-winged blackbird host nest?» (en inglés). Animal Behaviour (Association for the Study of Animal Behaviour) 55: p. 1181-1189. http://www3.amherst.edu/~edclotfelter/research_page/research_pdfs/Clotfelter_Animal_Behaviour_1998.pdf. Consultado el 10 de agosto de 2011.
- ↑ a b Berger, Andrew J. (marzo de 1951). «The cowbird and certain host species in Michigan» (en inglés). The Wilson Bulletin (Wilson Ornithological Society) 63 (1): p. 26-34. http://64.106.42.23/sora/Wilson/v063n01/p0026-p0034.pdf. Consultado el 9 de octubre de 2011.
- ↑ Linz, George M.; Bolin, Susan B. (marzo de 1982). «Incidence of brown-headed cowbird parasitism on red-winged blackbirds» (en inglés). The Wilson Bulletin (Wilson Ornithological Society) 94 (1): p. 93-95. http://www.jstor.org/stable/4161589. Consultado el 5 de octubre de 2011.
- ↑ a b Clotfelter, Ethan D.; Yasukawa, Ken (noviembre de 1999). «The effect of aggregated nesting on red-winged blackbird nest success and brood parasitism by brown-headed cowbirds» (en inglés). The Condor (Cooper Ornithological Society) 101 (4): p. 729-736. http://www.jstor.org/stable/1370059. Consultado el 9 de agosto de 2011.
- ↑ a b c d Rodenhouse, Nicholas L.; Best, Louis B.; O'Connor, Raymond J.; Bollinger, Eric K. (1992). «Effects of temperate agriculture on neotropical migrant landbirds» (en inglés). General Technical Report RM-229 (Fort Collins (Colorado): Departamento de Agricultura de Estados Unidos, Servicio Forestal, Rocky Mountain Forest and Range Experiment Station): p. 280-295. http://www.fs.fed.us/rm/pubs_rm/rm_gtr229/rm_gtr229_280_295.pdf. Consultado el 4 de abril de 2011.
- ↑ Besser, Jerome F.; Brady, Daniel J. (1986). «Bird damage to ripening field corn increases in the United States from 1971 to 1981» (en inglés). Fish and Wildlife Leaflet 7 (Washington D. C.: Departamento del Interior de los Estados Unidos, Servicio de Pesca y Vida Silvestre). http://www.dtic.mil/cgi-bin/GetTRDoc?AD=ADA323357&Location=U2&doc=GetTRDoc.pdf. Consultado el 9 de agosto de 2011.
- ↑ Bendell, Barry E.; Weatherhead, Patrick J.; Stewart, Robin K. (1981). «The impact of predation by red-winged blackbirds on European corn borer populations» (en inglés). Canadian Journal of Zoology (NRC Research Press) 59 (8): p. 1535-1538. http://www.nrcresearchpress.com/doi/abs/10.1139/z81-208. Consultado el 9 de agosto de 2011.
Enlaces externos
 Wikimedia Commons alberga contenido multimedia sobre Agelaius phoeniceus. Commons
Wikimedia Commons alberga contenido multimedia sobre Agelaius phoeniceus. Commons Wikiespecies tiene un artículo sobre Agelaius phoeniceus. Wikispecies
Wikiespecies tiene un artículo sobre Agelaius phoeniceus. Wikispecies
Categorías:- Pájaros cantores (Passeriformes) no amenazados
- Agelaius
- Aves migratorias del hemisferio occidental






















Wikimedia foundation. 2010.