- Visión de las aves
-
 Con ojos orientados hacia adelante el pigargo americano o águila calva tiene un amplio campo de visión binocular.
Con ojos orientados hacia adelante el pigargo americano o águila calva tiene un amplio campo de visión binocular.
La visión es el más importante de los sentidos de las aves, dado que es esencial para un vuelo seguro, y este grupo tiene un número de adaptaciones que permiten una agudeza visual superior a la de otros grupos de vertebrados. Para resaltar esto, por ejemplo, se ha descrito a la paloma como "dos ojos con alas".[1] El ojo de las aves se parece al de los reptiles actuales, pero tiene un mejor posicionamiento de los lentes, una característica compartida con los mamíferos. Las aves tienen los ojos más grandes en relación al tamaño corporal entre todo el reino animal, y el movimiento dentro de la cavidad ósea ocular es limitado por esa causa.[1] Adicionalmente a los dos párpados que siempre se encuentran en otros vertebrados, el ojo es protegido por una tercera membrana, móvil y transparente. La anatomía interna del ojo es similar a la de otros vertebrados, pero tiene una estructura, el pecten, única de las aves.
Las aves , como los peces, anfibio y reptiles, tienen cuatro tipos de receptores de color en el ojo. La mayoría de los mamíferos tienen dos tipos, aunque los primates tienen tres. Esto les da a las aves la habilidad de percibir no solo la luz en el espectro visible, sino también en el ultravioleta, y otras adaptaciones permiten la detección de luz polarizada. Las aves tienen proporcionalmente más receptores de luz en la retina que los mamíferos, y más conexiones nerviosas entre los fotorreceptores y el cerebro.
Algunos grupos de aves tienen modificaciones específicas para su sistema visual ligadas a sus modos de vida. Las aves de presa tienen una alta densidad de receptores y otras adaptaciones para maximizar la agudeza visual. La posición de sus ojos les dan buena visión binocular permitiéndoles una adecuada apreciación de las distancias. Las especies nocturnas tienen ojos tubulares, con poco número de detectores de color, pero una alta densidad de células bastoncillos, las que detectan mejor la luz menos intensa. Los charranes, gaviotas y albatros están entre las aves marinas que tienen gotas de aceite rojas y amarillas en los receptores de color para mejorar la visión a distancia, especialmente en condiciones brumosas.
Contenido
Anatomía extraocular
El parecido mayor del ojo de un ave es con el de los reptiles. A diferencia del ojo de mamíferos, no es esférico, y la forma aplanada permite que sea enfocado un mayor campo visual. Un círculo de placas óseas, el anillo esclerótico, rodea el ojo y lo mantiene rígido, pero una mejora sobre el ojo de los reptiles, que también se encuentra en mamíferos, es que el lente es empujado más hacia adelante, incrementando el tamaño de la imagen en la retina.[2]
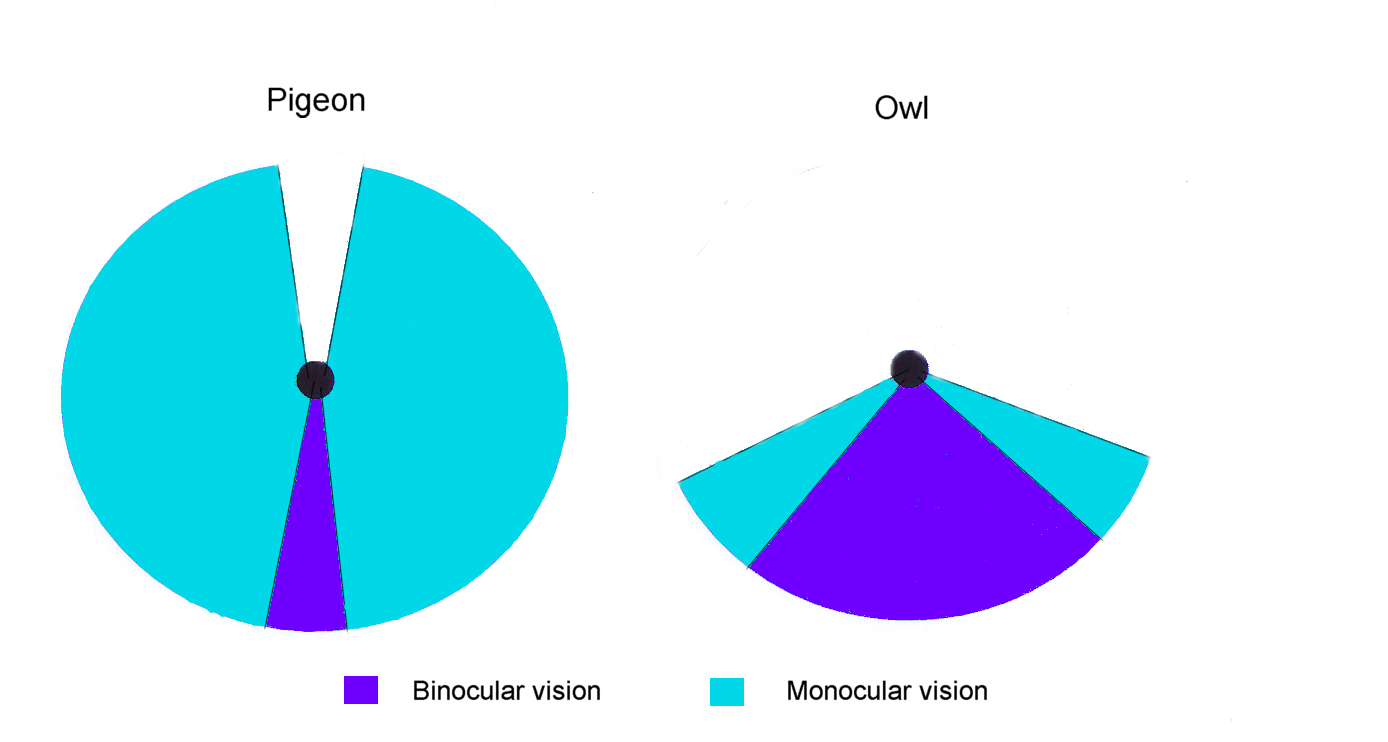
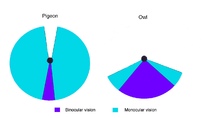 Campos visuales de un búho y de una paloma.
Campos visuales de un búho y de una paloma.La mayoría de las aves no pueden mover sus ojos, aunque hay excepciones, como el Cormorán Grande.[3] Las aves con ojos a los lados de la cabeza tienen un amplio campo visual, útil para la detección de predadores, mientras que aquellos con ojos al frente, como los búhos, tienen visión binocular y pueden estimar las distancias mientras cazan.[4] La Chocha Americana probablemente tiene el más amplio campo visual entre todas las aves, 360o en el plano horizontal, y 180o en el plano vertical.[5]
Los párpados de un ave no se usan en pestañear para la lubricación. En vez de los párpados, la función lubricadora la realiza la membrana nictitante, un tercer párpado oculto que se desliza horizontalmente por el ojo como un limpiaparabrisas.[6] La membrana nictitante también cubre el ojo y actúa como un lente de contacto en muchas aves acuáticas cuando están bajo el agua.[7] Cuando duermen el párpado inferior sube para cubrir el ojo en la mayoría de las aves, con la excepción del Búho Americano en el cual el párpado superior es móvil.[8]
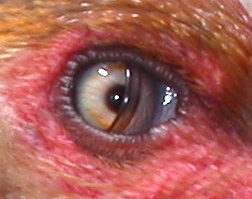
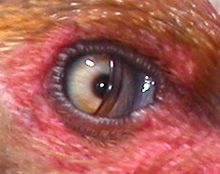 La membrana nictitante de una gallina.
La membrana nictitante de una gallina.El ojo es también limpiado por las secreciones de la glándula lagrimal y protegida por una sustancia oleosa segregada por la glándula de Harderian cubriendo la córnea y previniendo la sequedad. El ojo de un ave comparado con el tamaño del cuerpo es más grande que en cualquier otro grupo animal, aunque la mayor parte de éste está envuelto por el cráneo. El avestruz tiene el ojo más grande que en cualquier otro vertebrado terrestre, con una longitud axial de 50 mm, el doble que la del ojo humano.[1]
El tamaño del ojo de un ave está en relación aproximada con la masa corporal. Un estudio de cinco órdenes (loros, palomas, petreles, rapaces y búhos) mostró que la masa del ojo es proporcional a la masa corporal, pero como se esperaría por sus hábitos y ecología visual, las rapaces y los búhos tienen ojos relativamente grandes para sus masas corporales.[9]
Estudios de comportamiento muestran que muchas especies de aves enfocan los objetos lejanos preferentemente con su campo de visión lateral y monocular (con un solo ojo), y orientan la cabeza para ver por un lado y maximizar la resolución visual. Para una paloma, la resolución es el doble más fina con la visión monocular de lado que con la visión binocular frontal, mientras que en los humanos ocurre lo contrario.[1]
 El petirrojo europeo tiene ojos relativamente grandes, y comienza a cantar temprano en la mañana.
El petirrojo europeo tiene ojos relativamente grandes, y comienza a cantar temprano en la mañana.El desenvolvimiento del ojo a bajos niveles de luminosidad depende de la distancia entre el lente y la retina, y las aves pequeñas están efectivamente forzadas a ser diurnas debido a que sus ojos no son lo suficientemente grandes como para darles una adecuada visión nocturna. Aunque muchas especies migran durante la noche, a menudo ellas colisionan incluso con objetos brillantemente iluminados como faros y torres de perforación petrolera. Las aves de presa son diurnas porque, aunque sus ojos son grandes, ellas están optimizadas par obtener la máxima de resolución espacial en vez de la captación máxima de luz, así que sus ojos tampoco funciona bien con pobre iluminación.[10]
Muchas aves tienen una asimetría en la estructura del ojo que les permite mantener el horizonte y una parte significativa del suelo enfocados simultáneamente. El costo de esta adaptación es que ellas tienen miopía en la parte baja de su campo de visión.[1]
Las aves con ojos relativamente grandes comparados con sus masas corporales, como el colirrojo real y petirrojo europeo cantan más temprano al alba que otras aves del mismo tamaño y menor masa corporal. Sin embargo, si las aves tienen el mismo tamaño de ojo pero diferente masa corporal, la especie mayor canta más tarde que la menor. Esto puede ser debido a que el ave menor tiene que empezar la actividad diaria más temprano debido a la pérdida de peso nocturna.[11]
Las aves nocturnas tienen los ojos optimizados para la sensibilidad visual, con córneas grandes en relación a la longitud del ojo, mientras que las aves diurnas tienen ojos de mayor longitud en relación al diámetro de la córnea para darles mayor agudeza visual. El tipo de actividad de las especies extintas puede ser deducido de las mediciones del anillo esclerótico y la profundidad de la órbita ocular. Para poder hacer esta última medición, el fósil debe haber mantenido su forma tridimensional, así el patrón de actividad no puede determinarse con seguridad de especímenes aplastados como Archaeopteryx, el que tiene un anillo esclerótico completo pero no es posible hacerle mediciones de la profundidad de la órbita.[12]
Anatomía del ojo
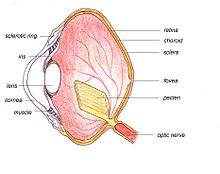 Anatomía del ojo de ave.
Anatomía del ojo de ave.Las estructuras principales del ojo de ave son similares a aquellas de otros vertebrados. La capa exterior del ojo consiste de la córnea transparente al frente, y dos capas de esclerótica —una capa de fibra de colágeno dura y blanca que recubre el resto del ojo, sosteniendo y protegiendo su integridad. El ojo está dividido internamente por el lente en dos segmento principales: el segmento anterior y el segmento posterior. La cámara anterior está llena de un fluido acuoso llamado humor acuoso, y la cámara posterior contiene el humor vítreo, una sustancia transparente de apariencia gelatinosa.
El lente es un cuerpo transparente y convexo con una capa exterior más dura y una capa interior más blanda. Éste enfoca la luz en la retina. Su forma puede ser alterada por medio de músculos ciliares que están adheridos al ojo por medio de las fibras zonulares, En adición a estos músculos, algunas aves también tienen un segundo juego de músculos, los músculos de Crampton, que pueden cambiar la forma de la córnea, lo que da a las aves una mayor amplitud de acomodación de lo que es posible en los mamíferos. El iris es un diafragma coloreado, operado por músculos delante del lente el cual controla la cantidad de luz que entra al ojo. Al centro del iris está la pupila, el área circular variable a través de la cual la luz pasa al interior del ojo.[2]
La retina es una estructura curva relativamente lisa de multiples capas, que contiene las células fotosensibles bastoncillos y conos con neuronas asociadas y vasos sanguíneos. La densidad de fotorreceptores es crítica en la determinación de la máxima agudeza visual asequible. Los humanos tienen cerca de 200.000 receptores por mm2, pero el gorrión doméstico tiene 400.000 y el busardo ratonero 1.000.000. Los fotorreceptores no están todos conectados individualmente al nervio óptico, y la relación de ganglios neurales respecto a receptores es importante en la determinación de la resolución. Ésta es muy alta para la aves; la lavandera blanca tiene 100.000 células gangionares para 120.000 fotorreceptores.[2]
Los bastoncillos son más sensibles a la luz, pero no dan información de color, mientras que los menos sensibles conos permiten la visión de color. En las aves diurnas, el 80% de los receptores pueden ser conos (90% en algunos vencejos) mientras que los búhos nocturnos casi todos los receptores son bastoncillos. Como en otros vertebrados excepto mamíferos placentados, algunos de los conos pueden ser de doble estructura. Éstos pueden sumar hasta 50% de todos los conos en algunas especies.[13]
Hacia el centro de la retina está la fóvea la cual tiene una densidad más grande de receptores y es el área de mayor agudeza visual delantera, es decir detección más detallada y clara de objetos. En 54% de las aves, incluidas aves de presa, martines pescadores, colibríes y golondrinas, existe una segunda fovea para una mejor visión de los laterales. El nervio óptico es un amasijo de fibras nerviosas que llevan mensajes desde el ojo hacia las partes relevantes del cerebro y viceversa. Como en los mamíferos, las aves tienen un pequeño punto ciego sin fotorreceptores dentro del disco óptico, bajo el cual el nervio óptico y los vasos sanguíneos se juntan en el ojo.[2]
El pecten es un cuerpo pobremente comprendido, de tejido plegado que se proyecta desde la retina. Está suministrado con vasos sanguíneos y parece mantener la retina suministrada de nutrientes,[1] y puede también apantallar la retina contra la luz deslumbrante o ayudar en la detección objetos en movimiento.[2]
El coroide es una capa situada detrás de la retina que contiene muchas pequeñas arterias y venas. Éstas respectivamente alimentan con sangre arterial y drenan la sangre venosa de la retina. El coroide contiene melanina, un pigmento que le da al interior del ojo su coloración oscura, lo que evita los reflejos trastornantes.
Percepción de la luz
Existen dos tipos de receptores de luz en el ojo de un ave bastoncillo y conos. Los bastoncillos, los que contienen el pigmento visual rodopsina son mejores para la visión nocturna porque son sensibles a pequeñas cantidades de luz. Los conos detectan luz de colores (o longitudes de ondas) específicos, así que son más importantes para animales orientados al color, tales como la mayoría de las aves.[16] La mayoría de las aves son tetracromáticas, poseyendo células conos sensibles al ultravioleta (UV) en el ojo, además de las sensibles al rojo, al verde y al azul,[17] pero las palomas tienen un pigmento adicional y por lo tanto son pentacromáticas.[18]
Los cuatro pigmentos de conos espectralmente diferentes se derivan de la proteína opsina, ligada a una pequeña molécula llamada retinal, la cual está estrechamente relacionada con la vitamina A. Cuando el pigmento absorbe luz el retinal cambia de forma y altera el potencial de membrana de la célula cono afectando las neuronas en la capa ganglionar de la retina. Cada neurona en la capa ganglionar puede poseer información de un número de células fotorreceptoras, y puede en consecuencia permitir disparar un impulso nervioso para transmitir información a lo largo del nervio óptico para un posterior procesamiento en centros visuales especializados en el cerebro. A luz más intensa, más fotones son absorbidos por pigmentos visuales, y mayor es la excitación de cada cono, y más brillante es la apreciación de esa luz.[16]
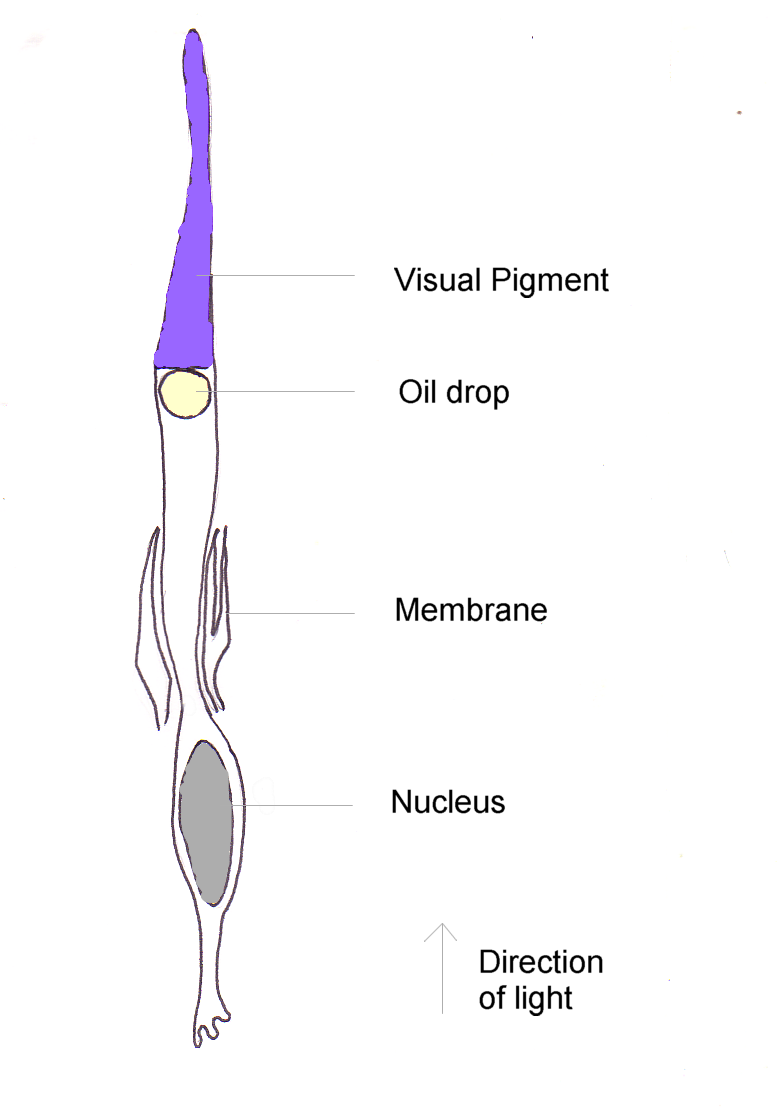
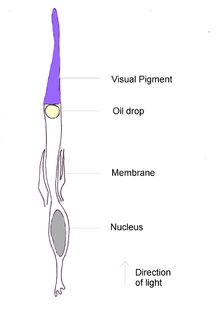 Diagrama de una célula cono de ave.
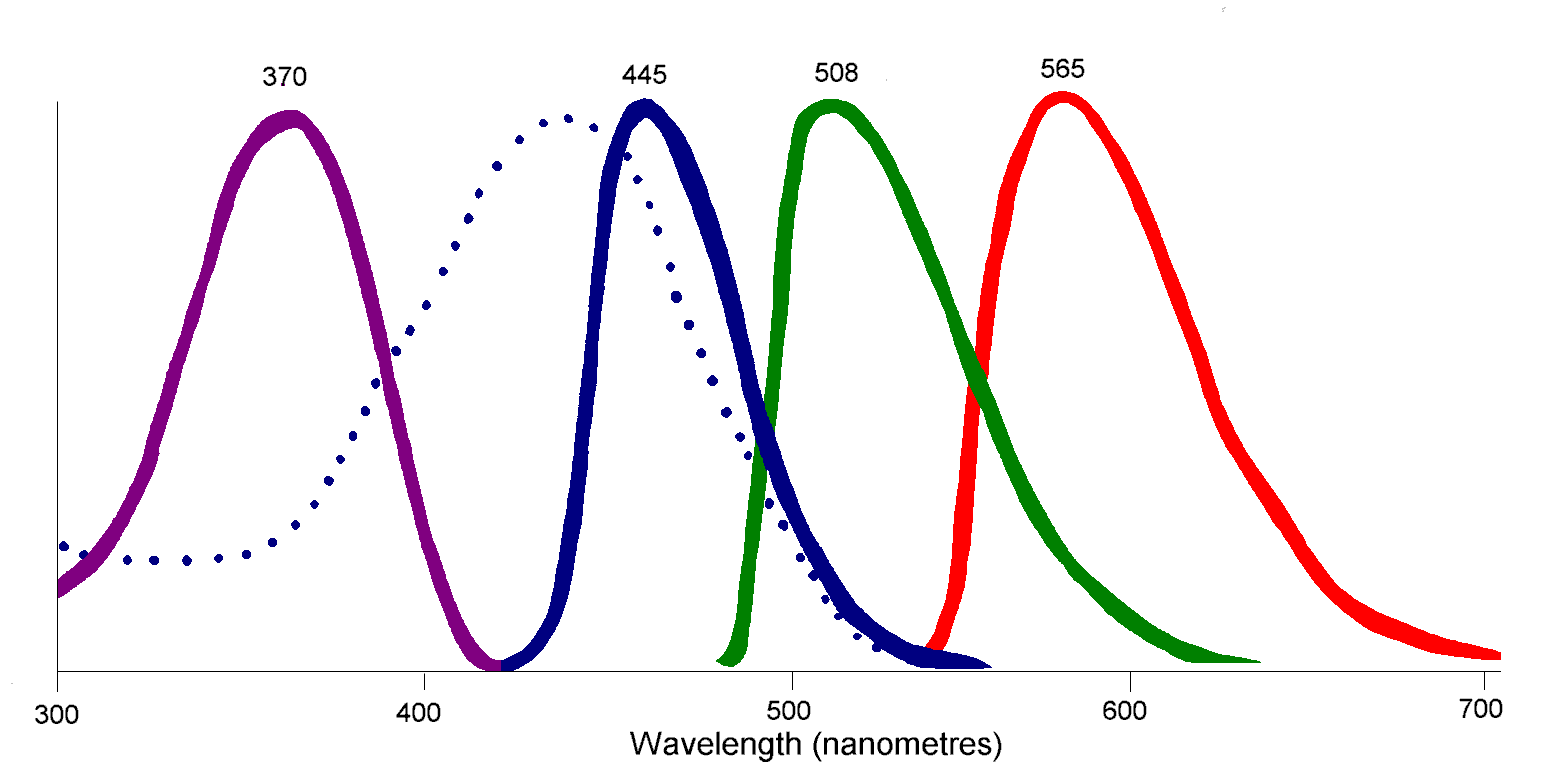
Diagrama de una célula cono de ave.El pigmento de conos más abundante y en alto grado es la forma de iodopsina de longitud de onda larga, la que absorbe cerca de los 570 nm. Esta es aproximadamente la región del espectro ocupada por los pigmentos sensibles al rojo y al verde en la retina de los primates, y este pigmento visual domina la sensibilidad al color en las aves.[18] En los pingüinos, este pigmento parece tener desplazado su pico de absorción a 543 nm, presumiblemente una adaptación al ambiente acuático azul.[19]
La información enviada por un único cono es limitada: por si sola la célula no puede decir al cerebro cuál longitud de onda causó la excitación. Un pigmento visual puede absorber dos longitudes de onda de igual forma, pero aunque sus fotones respectivos contienen energías diferentes, el cono no puede discriminarlas, porque ambos causan que el retinal cambie de forma y por lo tanto dispare el mismo impulso. Para que el cerebro aprecie el color, debe comparar las respuestas de dos o más clases de conos conteniendo diferentes pigmentos visuales, así los cuatro pigmentos en las aves brindan una discriminación incrementada.[16]
Cada cono de un ave o un reptil contiene una pequeña gota de aceite coloreada; éstas no existen en mamíferos. Las gotitas, que contienen concentraciones altas de carotenoides , están ubicadas de forma que la luz les pase a través antes de alcanzar el pigmento visual. Ellas actúan como filtros, eliminando algunas longitudes de ondas y estrechando el espectro de absorción de los pigmentos. Esto reduce la superposición de respuestas entre los pigmentos e incrementa el número de colores que un ave puede discernir.[16] Se han identificado seis tipos de gotitas de aceite de los conos; cinco de estos tienen mezclas de carotenoides que absorben a diferentes longitudes de ondas e intensidades, y el sexto tipo no tiene pigmentos.[20]
Los colores y distribuciones de las gotitas de aceite en la retina varían considerablemente entre las especies, y es más dependiente del nicho ecológico utilizado (cazador, pescador, herbívoro) que de las relaciones genéticas. Como ejemplos, cazadores diurnos como la golondrina común y las aves de presa tienen pocas gotitas coloreadas, mientras que el charrán común, pescador de superficie, tiene un gran número de gotitas rojas y amarillas en la retina dorsal. La evidencia sugiere que las gotitas de aceite responden a la selección natural en forma más rápida que los pigmentos visuales de los conos.[18]
Las aves canoras migratorias usan los campos magnéticos de la Tierra, el patrón de las estrellas y el Sol, y los patrones de luz polarizada para determinar su dirección migratoria. Un estudio estadounidense mostró que el migratorio gorrión sabanero común usa la luz polarizada de un área del cielo cercana al horizonte para recalibrar su sistema de orientación magnético durante la salida y la puesta del Sol. Esto sugiere que los patrones de polarización de la luz del cielo son la referencia de calibración primaria para todas la aves canoras migratorias.[21] Sin embargo, parece que las aves pueden estar respondiendo a indicadores secundarios del ángulo de polarización, y no pueden ser realmente capaces de detectar directamente la dirección de polarización en ausencia de estos indicios.[22]
Ultravioleta
 El cernícalo común puede detectar los rastros de orina que reflejan el ultravioleta dejados por sus presas, los campañoles.
El cernícalo común puede detectar los rastros de orina que reflejan el ultravioleta dejados por sus presas, los campañoles.Las aves pueden percibir la luz ultravioleta, la cual está involucrada en el cortejo. Muchas aves muestran patrones ultravioletas en los plumajes que son invisibles para el ojo humano. Los herrerillos europeos machos tienen un parche en la coronilla que refleja el ultravioleta el cual se exhibe en el cortejo por posturas y elevando las plumas de la nuca.[23] El azulejón o picogrueso azul macho que tenga el plumaje azul más brillante y corrido hacia el ultravioleta mantiene territorios más extensos con abundantes presas, y alimenta sus crías más frecuentemente que lo que consiguen hacerlo otros machos.[16]
La apariencia del pico es importante en la interacción del mirlo común. Aunque el componente UV no parece importante en interacciones entre machos que mantienen territorios, donde el grado de naranja es el factor principal, la hembra responde más fuertemente a machos con picos buenos reflectantes del UV.[24]
Un receptor de UV puede darle a un animal una ventaja en la búsqueda de comida. Las superficies cerosas de muchas frutas y bayas reflejan la luz UV que podría anunciar su presencia.[16] Los cernícalos comunes son capaces de localizar visualmente los rastros de ratas de agua o campañoles. Estos pequeños roedores dejan rastros de esencias de orina y heces que reflejan la luz UV, haciéndolas visibles a los cernícalos, particularmente en la primavera antes de que los rastros sean cubiertos por la vegetación.[25]
Percepción del movimiento
 Un cernícalo común necesita para cazar una imagen visual segura.
Un cernícalo común necesita para cazar una imagen visual segura.Las aves tiene mejor resolución de movimientos rápidos que los humanos, para quienes aleteos a una velocidad mayor que 50 Hz aparecen como movimiento continuo. Los humanos no pueden por lo tanto distinguir los parpadeos individuales de una lámpara de luz fluorescente que oscila a 60 Hz, pero los periquitos comunes y las gallinas tienen un umbral de resolución de movimiento de más de 100 Hz. Un gavilán de Cooper puede perseguir presas ágiles a través de bosques y evitar ramas y otros objetos a alta velocidad; éstos objetos para los humanos aparecerían como un borrón.[5]
Las aves pueden también detectar objetos de movimiento lento. El movimiento del sol a través del cielo es imperceptible para los humanos, pero detectable para las aves. La habilidad de detectar estos movimientos les permite a las aves migratorias orientarse correctamente.[5]
Para obtener imágenes seguras mientras vuelan o cuando están posadas sobre una rama oscilante, las aves mantienen la cabeza tan estable como es posible con reflejos compensatorios. El mantenimiento de imágenes estables es especialmente relevante para las aves de presa.[5]
Percepción de campos magnéticos
La percepción de campos magnéticos por aves migratorias se ha sugerido que es dependiente de la luz.[26] Las aves mueven la cabeza para detectar la orientación de los campos magnéticos.[27] Estudios sobre las rutas nerviosas han sugerido que las aves son capaces de "ver" los campos magnéticos.[28]
Aves de presa diurnas
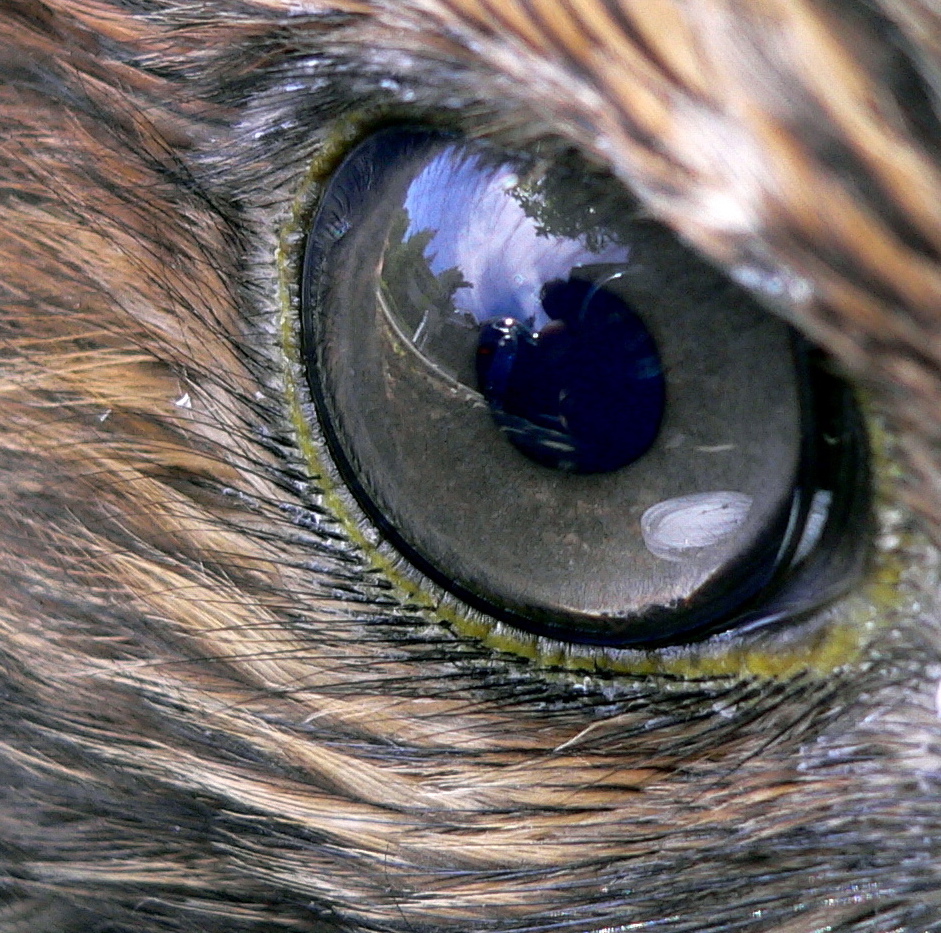
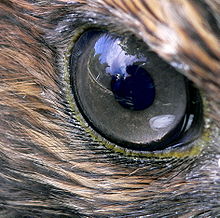 Ojo de un halcón. Tener "vista de águila" es una expresión para describir agudeza visual.
Ojo de un halcón. Tener "vista de águila" es una expresión para describir agudeza visual.La habilidad visual de las aves de presa es legendaria, y la agudeza de su visión es debida a una variedad de factores. Las rapaces tienen ojos grandes para su tamaño, 1,4 veces mayores que el promedio de las ave de su mismo peso,[9] y el ojo tiene forma de tubo para producir una imagen más grande en la retina. La retina tiene un gran número de receptores por milímetro cuadrado, lo que determina el grado de agudeza visual. Cuanto más receptores tenga un animal, mayor es la habilidad de distinguir objetos individuales a una distancia, especialmente cuando, como en las rapaces, cada receptor está típicamente unido a una sola célula ganglionar.[1]
Mucahs rapaces tienen foveas con más bastoncillos y conos que la fovea humana (65.000 mm2 en el Cernícalo Americano, 38.000 en humanos) y esto provee a estas aves con espectacular visión a larga distancia. La fovea misma puede ser de forma de lente, incrementando adicionalmente la densidad efectiva de receptores. Esta combinación de factores le da a los Buteo (busardos o gavilanes) la visión a distancia 6 a 8 veces mejor que la de humanos.
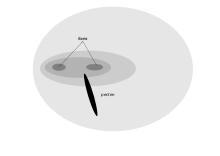
Los ojos dispuestos al frente de la cara de un ave de presa le dan visión binocular, la cual está asistida por una fovea doble.[2] Las adaptaciones de las rapaces para una resolución visual óptima (un cernícalo americano puede ver un insecto de 2–mm desde la cima de un árbol de 18–m) tiene una desventaja en que su visión es pobre a niveles de luz pobre, y deben descansar a la noche.[1] Las rapaces pueden tener que perseguir sus presas móviles en la parte más baja de su campo visual, y por lo tanto no tienen la adaptación de miopía de campo bajo demostrada en muchas otras aves.[1] Las aves carroñeras como los buitres no necesitan tal agudeza visual, así un cóndor tiene una sola fovea con cerca de 35.000 receptores por mm2
Las rapaces carecen de las gotitas de aceite coloreado en los conos, y probablemente tienen la percepción de color más similar a los humanos, y carecen de la habilidad de detectar luz polarizada. Los plumajes generalmente castaños, grises y blancos de este grupo, y la ausencia de exhibición de colores en el cortejo sugieren que el color no es relativamente importante en estas aves.[2]
En la mayoría de las rapaces una prominente arruga y sus plumas se extienden sobre y frente al ojo. Esta "ceja" le da a las aves de presa su mirada distintiva. La arruga protege físicamente al ojo del viento, polvo y desechos y lo apantalla del exceso de luz brillante. El águila pescadora carece de esta arruga, aunque el arreglo de las plumas sobre el ojo le sirve con similar función; también posee plumas oscuras en el frente del ojo lo que probablemente le sirve para reducir el brillo desde la superficie del agua cuando el ave está a la caza de peces, que son parte principal de su dieta.[5]
Aves nocturnas
Los búhos tienen grandes ojos para su tamaño, 2,2 veces mayores que el promedio para aves de su mismo peso,[9] y ubicados al frente de la cabeza. Los ojos tienen una superposición de campos visuales de los ojos de un 50–70%, lo que da mejor visión binocular que la que tienen las aves de presa diurnas (cuya superposición es de 30–50%).[30] La retina del cárabo común tiene cerca de 56.000 bastoncillos sensibles a la luz por milímetro cuadrado; aunque las antiguas declaraciones de que podía ver en la parte infrarroja del espectro han sido desechadas.[31]
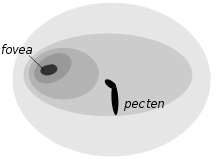 Cada retina de búho tiene una única fovea.[29]
Cada retina de búho tiene una única fovea.[29]La adaptaciones a la visión nocturna incluyen el gran tamaño del ojo, su forma tubular, gran número de bastoncillos dispuestos apretadamente, y la ausencia de conos, dado que la visión de color es innecesaria en la noche. Existen pocas gotas coloreadas, las cuales reducirían la intensidad de la luz, pero la retina contiene una capa reflectante, el tapetum lucidum. Éste incrementa la cantidad de luz que cada célula fotosensible recibe, permitiendo al ave ver mejor en condiciones de poca luz.[2] Los búhos tienen normalmente sólo una fovea, y ésta está poco desarrollada excepto en cazadoras diurnas como el búho campestre.[30]
Otras aves además de los búhos tienen buena visión nocturna, como el milano murcielaguero, los podargos y los añaperos. Algunas aves anidan en lo profundo de sistemas cavernarios los que son demasiado oscuros para la visión, y encuentran su camino al nido con un simple sistema de ecolocalización. El guácharo es la única ave nocturna que ecolocaliza,[32] pero varias Aerodramus (salanganas) también utilizan esta técnica, con una especie, la salangana de las Cook, que también usa la ecolocalización fuera de sus cuevas.[33] [34]
Aves acuáticas
 Los charranes tienen gotitas de aceite en las células cono del ojo para mejorar la visión a distancia.
Los charranes tienen gotitas de aceite en las células cono del ojo para mejorar la visión a distancia.Las ave marinas como los charranes y gaviotass que se comen de la superficie o se zambullen por el alimento tienen gotitas de aceite rojas en los conos de de sus retinas. Esto les mejora el contraste y agudiza su visión a distancia, especialmente en condiciones brumosas.[2] Birds that have to look through an air/water interface have more deeply coloured carotenoid pigments in the oil drops than other species.[18] Esto las ayuda a localizar cardúmenes de peces, aunque no hay certeza si es que están viendo el fitoplancton del cual los peces se alimentan, u otras aves alimentándose.[35]
Las aves que persiguen peces bajo el agua como las alcas y colimbos tienen muchas menos gotitas de aceite rojas,[2] pero tienen lentes especiales flexibles y usan la membrana nictitante como una lente adicional. Esto les permite una mayor acomodación óptica para una buena visión en el aire y en el agua.[7] Los cormoranes tienen una amplitud de acomodación visual mayor, hasta 50 dioptrías, que cualquier otra ave, pero los martines pescadores están considerados como los de mejor visión en conjunto en ambos medios, aire y agua.[2]
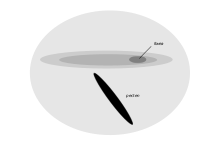 Cada retina de pardela pichoneta tiene una fovea y una banda alargada de alta densidad de receptores.[29]
Cada retina de pardela pichoneta tiene una fovea y una banda alargada de alta densidad de receptores.[29]Las aves marinas procelariformes (petreles y pardelas), las cuales vienen a la costa sólo a criar y permanecen la mayor parte errando cerca de la superficie de los océanos, tienen un área de sensibilidad visual larga y estrecha en la retina.[1] Esta región, el área de células gigantes, ha sido encontrada en especies como pardela pichoneta, petrel de Kerguelen, pardela capirotada, pato-petrel piquiancho y potoyunco común. Se caracteriza por la presencia de células ganglionares que están ordenadas regularmente y son más grandes que las halladas en el resto de la retina, y morfológicamente se parecen a las de la retina de gatos. La localización y morfología celular de esta novedosa área sugiere una función en la detección de objetos en un campo binocular pequeño proyectado debajo y alrededor del pico. No tiene que ver primariamente con alta resolución espacial, pero puede asistir en la detección de presas cerca de la superficie dado que estas aves vuelan a muy poca distancia de ésta.[36]
La pardela pichoneta, como muchas otras aves marinas, visitan sus colonias de cría durante la noche para reducir los riesgos de ataque por depredadores aéreos. Dos aspectos de su estructura óptica sugieren que el ojo de esta especie está adaptado a la visión nocturna. En el ojo de la pardela el lente hace la mayor parte de la refracción de luz necesaria para producir una imagen enfocada en la retina. La córnea, la capa exterior que cubre el frente del ojo, es relativamente aplanada y por lo tanto de bajo poder refractivo. En un ave diurna como la paloma, ocurre lo contrario; la córnea está altamente curvada y es el principal componente refractivo. La relación de la refracción producida por el lente respecto a la producida por la córnea es de 1,6 para la pardela y de 0,4 para la paloma; este índice en la pardela es consistente con el que está presente en un abanico diverso de aves y mamíferos nocturnos.[37]
La longitud de enfoque más corta de los ojos de las pardelas les da una imagen menor pero más brillante que la que obtienen las palomas, así es que estas últimas tienen una visión diurna más aguda. Aunque la pardela pichoneta tiene adaptaciones para la visión nocturna, el efecto es pequeño, y es probable que estas aves también usan el olfato y la audición para localizar sus nidos.[37]
Se solía pensar que los pingüinos eran miopes en tierra. Aunque la córnea es aplanada y adaptada para la visión subacuática, el lente es muy fuerte y puede compensar el enfoque reducido de la córnea cuando el ave sale del agua.[30] Casi la solución contraria es usada por la Serreta Capuchona la cual puede combar el lente a través del iris cuando está sumergida.[30]
Notas
- ↑ a b c d e f g h i j Güntürkün, Onur, "Structure and functions of the eye" in Sturkie (1998) 1–18
- ↑ a b c d e f g h i j k Sinclair (1985) 88–100
- ↑ White, Craig R.; Norman Day, Patrick J. Butler, Graham R. Martin (July 2007). «Vision and Foraging in Cormorants: More like Herons than Hawks?» (PDF). PLoS ONE 2 (7): pp. e639. doi:. PMID 17653266. http://eprints.bham.ac.uk/55/1/martin.pdf.
- ↑ Martin, Graham R.; Gadi Katzir (1999). «Visual fields in short-toed eagles, Circaetus gallicus (Accipitridae), and the function of binocularity in birds». Brain, Behaviour and Evolution 53 (2): pp. 55–66. doi:. PMID 9933782.
- ↑ a b c d e Jones, Michael P; Pierce, Kenneth E; Ward, Daniel (April 2007). «Avian vision: a review of form and function with special consideration to birds of prey» (PDF). Journal of Exotic Pet Medicine 16 (2): pp. 69–87. doi:. http://www.csulb.edu/~efernand/visualecol/Avian%20vision.pdf.
- ↑ Williams, David L.; Flach, Edmund (March 2003). «Symblepharon with aberrant protrusion of the nictitating membrane in the snowy owl (Nyctea scandiaca)» (PDF). Veterinary Ophthalmology 6 (1): pp. 11–13. doi:. PMID 12641836. http://www.davidlwilliams.org.uk/resources/file0016.pdf.
- ↑ a b Gill, Frank (1995). Ornithology. New York: WH Freeman and Co. ISBN 0-7167-2415-4. OCLC 30354617.
- ↑ The bird: its form and function. Henry Holt & Co, New York. 1906. pp. 214. http://www.archive.org/details/birditsformfunct00beeb.
- ↑ a b c Brooke, M. de L.; Hanley, S.; Laughlin, S. B. (February 1999). «The scaling of eye size with body mass in birds» (PDF). Proceeding of the Royal Society Biological Sciences 266: pp. 405–412. http://rspb.royalsocietypublishing.org/content/suppl/2009/02/12/269.1493.831.DC1/rspb20011941s01.pdf.
- ↑ Martin, Graham. "Producing the image" in Ziegler & Bischof (1993) 5–24
- ↑ Thomas, Robert J.; Székely, Tamás; Cuthill, Innes C.; Harper, David G. C.; Newson, Stuart E.; Frayling, Tim D.; Wallis, Paul D. (2002). «Eye size in birds and the timing of song at dawn» (PDF). Proceedings of the Royal society of London 269 (1493): pp. 831–837. doi:. PMID 1690967. http://www.pubmedcentral.nih.gov/picrender.fcgi?artid=1690967&blobtype=pdf.
- ↑ Hall, Margaret I. (junio 2008). «The anatomical relationships between the avian eye, orbit and sclerotic ring: implications for inferring activity patterns in extinct birds». Journal of Anatomy 212 (6): pp. 781–794. doi:.
- ↑ Nalbach Hans-Ortwin; Wolf-Oberhollenzer, Friedericke; Remy Monika. "Exploring the image" in Ziegler & Bischof (1993) 26–28
- ↑ El efecto de las gotitas de aceite coloreadas es desplazar el pico de absorción de cada pigmento. La línea de puntos muestra el pico de absorción del pigmento 445 sin desplazar. Los pigmentos 508 y 565 mostrarían un efecto similar, pero las absorciones no desplazadas de los ellos se omiten para mayor claridad.
- ↑ Basado en Goldsmith (2006).
- ↑ a b c d e f Goldsmith, Timothy H. (July 2006). «What birds see» (PDF). Scientific American: pp. 69–75. http://www.csulb.edu/labs/bcl/elab/avian%20vision_intro.pdf.
- ↑ Wilkie, Susan E.; Vissers, Peter M. A. M.; Das, Debipriya; Degrip, Willem J.; Bowmaker, James K.; Hunt, David M. (1998). «The molecular basis for UV vision in birds: spectral characteristics, cDNA sequence and retinal localization of the UV-sensitive visual pigment of the budgerigar (Melopsittacus undulatus)» (PDF). Biochemical Journal 330: pp. 541–47. PMID 9461554. http://www.pubmedcentral.nih.gov/picrender.fcgi?artid=1219171&blobtype=pdf.
- ↑ a b c d Varela, F. J.; Palacios, A. G.; Goldsmith T. M. "Color vision in birds" in Ziegler & Bischof (1993) 77–94
- ↑ Bowmaker, J. K.; Martin, G. R. (January 1985). «Visual pigments and oil droplets in the penguin, Spheniscus humbolti». Journal of Comparative Physiology 156 (1): pp. 71–77. doi:.
- ↑ Goldsmith, T. H.; Collins, J. S;. Licht, S. (1984). «The cone oil droplets of avian retinas». Vision Research. 24 (11): pp. 1661–1671. doi:. PMID 6533991.
- ↑ Muheim, Rachel; Phillips, John B. Åkesson, Susanne (August 2006). «Polarized light cues underlie compass calibration in migratory songbirds» (PDF). Science 313: pp. 837–839. doi:. PMID 16902138. http://www.angel.ekol.lu.se/~rachel/publications/JOrnithol%202007.pdf.
- ↑ Greenwood, Verity J.; Smith, Emma L.; Church, Stuart C.; Partridge, Julian C. (2003). «Behavioural investigation of polarisation sensitivity in the Japanese quail (Coturnix coturnix japonica) and the European starling (Sturnus vulgaris)». The Journal of Experimental Biology 206: pp. 3201–3210. doi:. PMID 12909701. http://jeb.biologists.org/cgi/content/full/206/18/3201.
- ↑ Andersson, S. (1998). «Ultraviolet sexual dimorphism and assortative mating in blue tits». Proceeding of the Royal Society B 265 (1395): pp. 445–50. doi:. http://beheco.oxfordjournals.org/cgi/content/full/15/5/805.
- ↑ Bright, Ashleigh.; Waas, Joseph R. (August 2002). «Effects of bill pigmentation and UV reflectance during territory establishment in blackbirds» (PDF). Animal Behaviour 64 (2): pp. 207–213. doi:. http://cber.bio.waikato.ac.nz/images/bbposter2.pdf.
- ↑ Viitala, Jussi; Erkki Korplmäki, Pälvl Palokangas & Minna Koivula (1995). «Attraction of kestrels to vole scent marks visible in ultraviolet light». Nature 373 (6513): pp. 425–27. doi:.
- ↑ «Night-vision brain area in migratory songbirds». PNAS 102: pp. 8339-8344. 2005. doi:.
- ↑ Mouritsen, H.; G.Feenders, M.Liedvogel, W.Kropp (2004). «Migratory Birds Use Head Scans to Detect the Direction of the Earth"s Magnetic Field». Current Biology 14 (21): pp. 1946-1949. doi:. http://www.seaturtle.org/PDF/Mouritsen_2004_CurrBiol.pdf.
- ↑ Heyers D, Manns M, Luksch H, Güntürkün O, Mouritsen H (2007). «A Visual Pathway Links Brain Structures Active during Magnetic Compass Orientation in Migratory Birds.». PLoS ONE 2 (9): pp. e937. doi:.
- ↑ a b c Diagrama esquemático de retina de ojo derecho, basado someramente en Sturkie (1998) 6
- ↑ a b c d Burton (1985) 44–48
- ↑ Hecht, Selig; Pirenne, Maurice Henri (1940). «The sensibility of the nocturnal long-eared owl in the spectrum» (Automatic PDF download). Journal of General Physiology 23: pp. 709–717. doi:. http://www.jgp.org/cgi/reprint/23/6/709.
- ↑ Cleere, Nigel; Nurney, David (1998). Nightjars: A Guide to the Nightjars, Frogmouths, Potoos, Oilbird and Owlet-nightjars of the World. Pica / Christopher Helm. pp. 7. ISBN 1873403488. OCLC 39882046.
- ↑ Fullard, J. H.; Barclay, R.M.R.; Thomas D.W. (1993). «Echolocation in free-flying Atiu Swiftlets (Aerodramus sawtelli)» (PDF). Biotropica 25: pp. 334–339. doi:. http://www.erin.utoronto.ca/%7Ew3full/reprints/1993FullBarcThomKopekaBiotropica.pdf. Retrieved 12 July 2008
- ↑ Konishi, M.; Knudsen E. I. (April 1979). «The oilbird: hearing and echolocation». Science 204: pp. 425–427. doi:. PMID 441731. Retrieved 12 July 2008
- ↑ Lythgoe, J. N. (1979). The Ecology of Vision. Oxford: Clarendon Press. pp. 180–183. ISBN 0198545290. OCLC 4804801.
- ↑ Hayes, Brian; Martin, Graham R.; Brooke, Michael de L. (1991). «Novel area serving binocular vision in the retinae of procellariiform seabirds». Brain, Behavior and Evolution 37 (2): pp. 79–84. doi:.
- ↑ a b Martin, Graham R.; Brooke, Michael de L. (1991). «The Eye of a Procellariiform Seabird, the Manx Shearwater, Puffinus puffinus: Visual Fields and Optical Structure». Brain, Behaviour and Evolution 37 (2): pp. 65–78. doi:.
- Este artículo fue creado a partir de la traducción del artículo Bird vision de la Wikipedia en inglés, bajo licencia Creative Commons Atribución Compartir Igual 3.0 y GFDL.
Bibliografía
- Burton, Robert (1985). Bird Behaviour. London: Granada Publishing. ISBN 0246124407.
- Sinclair, Sandra (1985). How Animals See: Other Visions of Our World. Beckenham, Kent: Croom Helm. ISBN 0709933363.
- Sturkie, P. D. (1998). Sturkie's Avian Physiology. 5th Edition. Academic Press, San Diego. ISBN 0-12-747605-9. OCLC 162128712 191850007 43947653.
- Ziegler, Harris Philip; Bischof, Hans-Joachim (eds.) (1993). Vision, Brain, and Behavior in Birds: A comparative review. MIT Press. ISBN 026224036X. OCLC 27727176.
Enlaces externos
 Wikimedia Commons alberga contenido multimedia sobre Visión de las aves. Commons
Wikimedia Commons alberga contenido multimedia sobre Visión de las aves. Commons











Wikimedia foundation. 2010.