- Iguanodon
-
 Iguanodon
IguanodonRango temporal: Cretácico inferior 
Clasificación científica Reino: Animalia Filo: Chordata Clase: Sauropsida Superorden: Dinosauria Orden: Ornithischia Suborden: Ornithopoda Infraorden: Iguanodontia Familia: Iguanodontidae Género: Iguanodon
Mantell, 1825Especies Sinonimia Iguanodon (gr. "diente de iguana") es un género de dinosaurios ornitópodos iguanodóntidos, que vivieron a principios período Cretácico, hace aproximadamente 130 a 120 millones de años (entre el Berriasiano y el Aptiano), en lo que hoy es Europa. Iguanodon está a medio camino entre los primitivos de los hipsilofodóntidos, bípedos y rápidos, y la culminación de los ornitópodos en los dinosaurios de pico de pato. Se han descrito muchas especies de Iguanodon, datadas desde el Kimeridgiano, a finales del Jurásico, al Cenomaniano del Cretácico Superior en Asia, Europa, y Norteamérica. Sin embargo, desde la primera década del siglo XXI se tiende a reconocer una única especie, Iguanodon bernissartensis, que vivió en el Cretácico Inferior de Europa. La principal característica de Iguanodon son sus grandes garras en los pulgares, que fueron utilizados posiblemente para defenderse de los depredadores.
Descubierto en 1822 y descrito tres años más tarde por el geólogo inglés Gideon Mantell, Iguanodon fue el segundo dinosaurio nombrado formalmente, después de Megalosaurus. Junto con Megalosaurus y Hylaeosaurus, es uno de los tres géneros usados originalmente para definir Dinosauria. Fue bautizado por Gideon Mantell con el nombre Iguanodon, el cual se deriva del término iguana y de la palabra griega odontos (diente), debido a que los dientes de Iguanodon son similares a los de la iguana. Fueron herbívoros grandes y robustos. Iguanodon es un miembro de Iguanodontia, junto con los hadrosáuridos. La taxonomía de este género continúa siendo un asunto de estudio mientras que se nombran nuevas especies o las de muchos años se reasignan a otros géneros.
La comprensión científica de Iguanodon ha evolucionado a través del tiempo, a medida que nueva información se obtiene de los fósiles. Los numerosos especímenes de este género, incluyendo los esqueletos casi completos a partir de dos niveles fosilíferos bien conocidos, han permitido que los investigadores se formulen hipótesis con respecto a muchos aspectos de la vida del animal, incluyendo la alimentación, el movimiento, y el comportamiento social. Como uno de los primeros dinosaurios bien conocidos por la ciencia, Iguanodon ha ocupado un lugar pequeño pero notable en la opinión pública sobre los dinosaurios, dado que sus representaciones artísticas han ido cambiando perceptiblemente en respuesta a las sucesivas interpretaciones de su anatomía.
Contenido
Anatomía
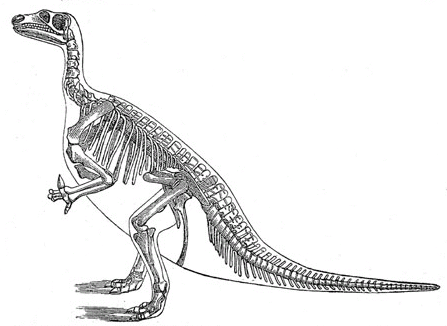
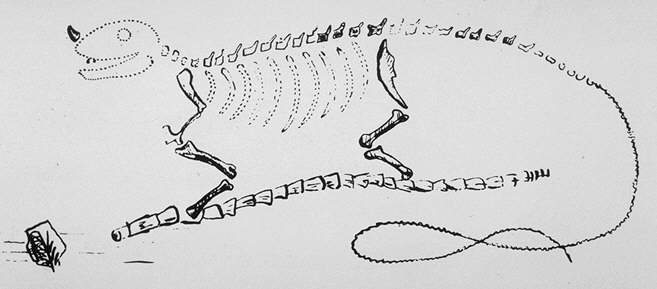
 Ilustración de la estructura ósea de un iguanodonte.
Ilustración de la estructura ósea de un iguanodonte.
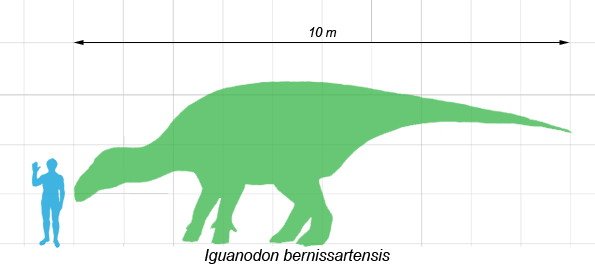
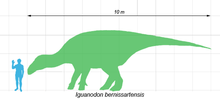 Iguanodon bernissartensis comparado con un ser humano.
Iguanodon bernissartensis comparado con un ser humano.Iguanodon fue un robusto herbívoro que podía alternar entre las marchas bípeda y cuadrúpeda.[1] Se ha estimado que los adultos de la especie mejor conocida, I. bernissartensis, pesaban un promedio de 3 toneladas,[2] y medían cerca de 10 metros de largo, pudiendo alcanzar en algunos casos los 13 metros.[3] Otras especies no eran tan grandes, el igualmente robusto I. dawsoni se estima en 8 metros de largo, y su contemporáneo más ligero, I. fittoni, en 6 metros.[4]
La espina dorsal y la cola estaban reforzadas y rigificadas por tendones osificados cilíndricos, que se omiten generalmente de los montajes y de los dibujos esqueléticos.[1] En general, su aspecto y estructura del cuerpo no difería de los posteriores hadrosáuridos.
Cráneo
 Cranéo de Iguanodon del Museo de Historia Natural de la Universidad de Oxford.
Cranéo de Iguanodon del Museo de Historia Natural de la Universidad de Oxford.Este género tenía un cráneo grande, alto pero estrecho, con un desdentado pico cubierto probablemente con queratina, y los dientes como los de una iguana, pero mucho más grandes y juntos en paquetes.[1] El cráneo estaba estructurado de tal manera que cuando cerraba la mandíbula, los maxilares (huesos del cráneo que sostienen los dientes superiores) se arqueaban hacia fuera. Esto causaría que las superficies más bajas de los dientes superiores frotaran contra la superficie superior de los dientes de la mandíbula, moliendo cualquier cosa entre ellos y proporcionando una acción equivalente a la masticación de los mamíferos.[5] Debido a que los dientes siempre eran sustituidos, el animal habría podido utilizar este mecanismo toda su vida, y podría comer material vegetal resistente.[6] Por otro lado, la parte frontal del hocico era desdentada, y estaría probablemete cubierta por un pico córneo, apto para arrancar ramitas y brotes,[7] ubicado sobre los huesos premaxilar (en el cráneo) y predentario (en la mandíbula), cada uno de los cuales formaba una punta arqueada rugosa que le servirían de base.[1]
Los iguanodontes poseían un robusto aparato masticador, dotado con la dentición típica de un reptil herbívoro.[8] El mismo Mantell notó que los restos con los que trabajaba eran distintos a los de cualquier reptil moderno, especialmente en la mandíbula inferior con una sínfisis en forma de cuchara, que comparó con el perezoso de dos dedos y el extinto perezoso terrestre Mylodon. Mantell sugirió que los iguanodontes habrían tenido una lengua prensil que podrían utilizar para recolectar el alimento,[9] como las jirafas actuales. Restos más completos que los estudiados por Mantell han demostrado que esto no podía ser correcto; así el hueso hioides de Iguanodon, en el que se insertan músculos de la lengua, tenía una estructura robusta, implicando una lengua muy musculosa, no prensil, incapaz para realizar los movimientos sugeridos. [10] La idea de la lengua prensil ha sido atribuida erróneamente a Dollo.[11]
Los iguanodontes tenían un único diente de reemplazo por cada pieza dentaria en uso, a diferencia de los hadrosáuridos, que tenían varias filas de dientes de reemplazo. Poseían 29 dientes en cada maxilar (58 dientes superiores), y 25 en cada dentario (50 dientes inferiores), mientras que en el premaxilar y el predentario, que formaban el «pico», no poseían ninguno. El mayor número de dientes superiores se compensa con una mayor longitud de los dientes de la mandíbula, lo que permitía la oclusión de las filas dentales en toda su longitud, necesaria para la masticación eficaz.[12] Debido ciertos detalles anatómicos y a la profunda inserción de los dientes desde el exterior de las mandíbulas se supone que, al igual que la mayoría de los demás ornitisquios, los iguanodontes tenían algún tipo de estructura similar a la mejilla, muscular o no, para conservar el alimento en la boca durante la masticación.[13] [14]
Extremidades
 Una mano de Iguanodon mostrada en Bruselas; con el dedo prensil extendido.
Una mano de Iguanodon mostrada en Bruselas; con el dedo prensil extendido.Las piernas eran poderosas, pero no aptas para la carrera, y tenían tres dedos en cada pie.
Los brazos eran largos, hasta 75% de la longitud de las piernas en I. bernissartensis y robustos,[3] con las manos poco flexibles construidas de modo que los tres dedos centrales (dedos II, III y IV) pudieran soportar el peso en posición cuadrúpeda.[1] El dedo meñique (dedo V) era alargado y flexible, y habría podido ser utilizado para manipular objetos y de ayuda en la recolección del alimento.[1]
Pulgares con garras
 Detalle de la mano de Iguanodon.
Detalle de la mano de Iguanodon. Abajo a la izquierda falange del pulgar encontrada en Maidstone en 1840 (interpretada entonces erroneamente como un cuerno nasal).
Abajo a la izquierda falange del pulgar encontrada en Maidstone en 1840 (interpretada entonces erroneamente como un cuerno nasal).Una de las características de los iguanodontes es el gran dedo pulgar (dedo I o pollex), cuya segunda falange formaba gran una garra cónica, muy desarrollada y probablemente dotada con una gruesa cubierta córnea.[15] Los pulgares estaban dirigidos hacia fuera, perpendiculares a los tres dedos centrales.
Este pulgar es tradicionalmente explicado como un elemento defensivo contra los predadores[7] [1] o contra otros iguanodontes,[3] aunque pudo usarlo también para partir semillas y frutas.[1] Se ha propuesto también la posibilidad de que la garra estuviera asociada a una glándula venenosa,[16] pero la hipótesis está en entredicho, ya que la falange ni es hueca,[3] ni existe ningún tipo de estría o conducto que pudiera conducir el veneno.[17]
En las reconstrucciones iniciales Mantell confundió la última falange del pulgar con un cuerno que ubicó sobre la nariz; así, con aspecto de rinoceronte, apareció representado Iguanodon en las primeras recreaciones, hasta que se hallaron los ejemplares completos en Bernissart y Dollo se dio cuenta del error, colocando el hueso en el lugar correcto de la mano: un pulgar modificado.[7] Ésta no sería la vez última que la garra modificada del pulgar de un dinosaurio era mal interpretada; Noasaurus, Baryonyx y Megaraptor son ejemplos desde la década de 1980, cuando las garras agrandadas del pulgar fueron puestas en el pie, como en los dromeosáuridos.
Historia
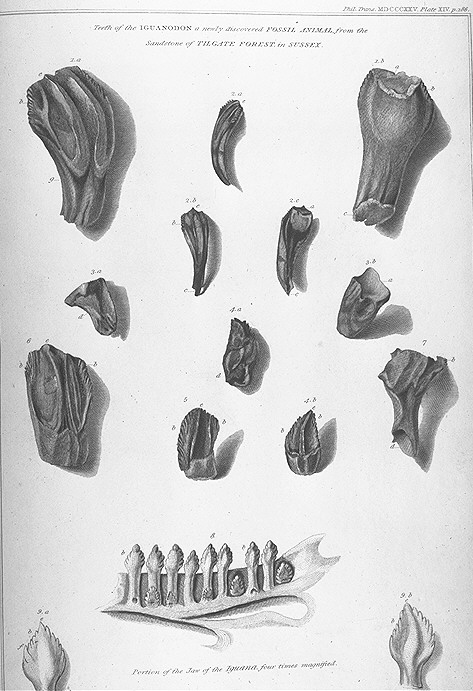
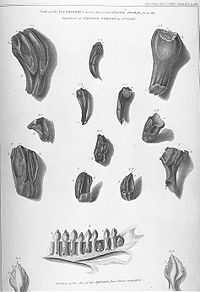 Ilustración de los fósiles de dientes de Iguanodon junto a los de una iguana moderna en el trabajo de Mantell de 1825.
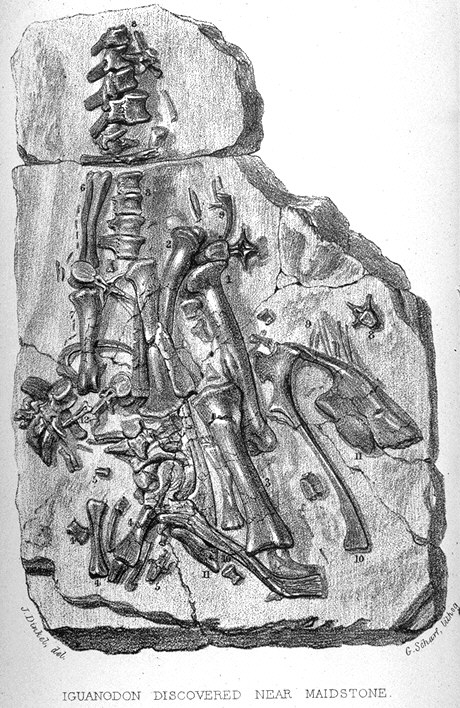
Ilustración de los fósiles de dientes de Iguanodon junto a los de una iguana moderna en el trabajo de Mantell de 1825.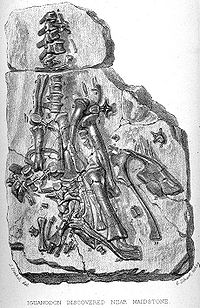 Restos encontrados en Maidstone en 1840.
Restos encontrados en Maidstone en 1840.El descubrimiento de Iguanodon ha sido acompañado a lo largo del tiempo por una popular leyenda. El primer rastro de un Iguanodon fue un diente fosilizado encontrado por la esposa de Gideon Mantell, Mary Ann Mantell, en los estratos del Bosque Tilgate (Cuckfield, Inglaterra), mientras su marido revisaba un paciente. Sin embargo, no hay evidencia de que Mantell llevara a su esposa con él mientras veía a sus pacientes. Además, admitió en 1851 que él mismo había encontrado los dientes.[18] Ninguno afirma que la otra historia es falsa[19] Sin importar las circunstancias exactas, inpeccionó el área en busca de más fósiles, y consultó a los expertos sobre fósiles de su tiempo en cuanto a qué clase de animal pudieron pertenecer los huesos. La mayor parte de los científicos, por ejemplo William Buckland y Georges Cuvier, pensaron que los dientes eran de pez o mamífero. Sin embargo, Samuel Stutchbury, un naturalista del Real Colegio de Cirujanos, reconoció que se asemejaban a los de una iguana, no obstante veinte veces de más grande.[2] Mantell no describió sus descubrimientos hasta 1825, cuando presentara un trabajo en la Royal Society de Londres.[8] [18] En el reconocimiento de la semejanza de los dientes a los de la iguana, Mantell nombró su nuevo género Iguanodon o "dientes de iguana", a partir de iguana y del griego odontos ("diente").[3] Basándose en una escala isometrica, estimó que la criatura habría alcanzado los 12 metros de largo.[8] La idea inicial fue llamarlo Iguanasaurus ("lagarto iguana "), pero su amigo William Daniel Conybeare le sugirió que ese nombre era más aplicable a la misma iguana y que mejor seria llamarlo Iguanoides ("como una iguana ") o Iguanodon.[20] [21] Tuvo el descuido de no asignar nombre a la especie para formar la apropiada nomenclatura binomial, así que el nombre fue suministrado en 1829 por Friedrich Holl: I. anglicum, que fue enmendado más adelante como I. anglicus.[22]
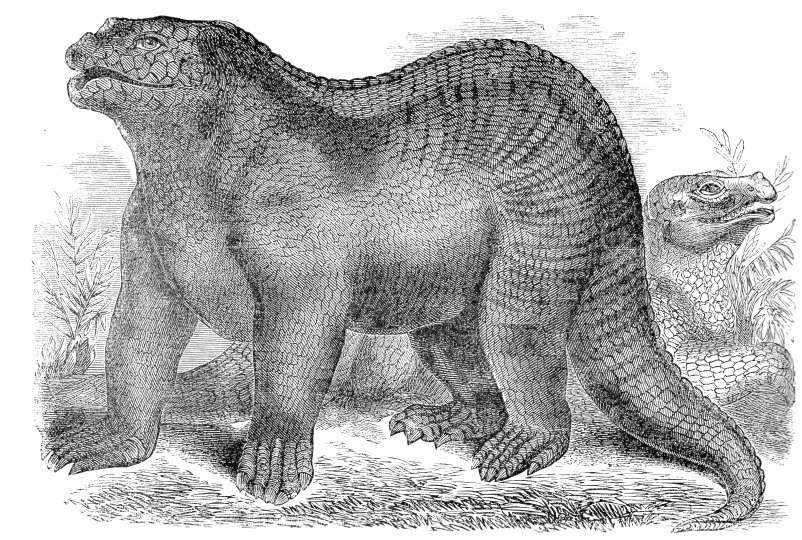
 Reconstrucción de Mantell de Iguanodon basada en los restos de Maidstone.
Reconstrucción de Mantell de Iguanodon basada en los restos de Maidstone.Años después se encontró un espécimen más completo de Iguanodon en una cantera de Maidstone, que Mantell adquirió e identificó basado en el diente que tenía. La losa de Maidstone permitió las primeras reconstrucciones esqueléticas e interpretaciones artísticas de un iguanodonte. El error más famoso fue la adición de un cuerno, también descubierto por la esposa de Mantell, sobre la nariz del dinosaurio.[23] El descubrimiento de muchos mejores especímenes de I. bernissartensis en 1878 reveló que el cuerno era en realidad un pulgar modificado, quizás usado para la defensa, a modo de puñal contra los depredadores de su ambiente. Dicho error puede observarse en las esculturas preparadas para la gran exposición de 1851 del Palacio de Cristal de Londres, conservadas actualmente como testimonio histórico. Para conmemorar el descubrimiento el distrito de Maidstone incorporó un Iguanodon como soporte en su escudo de armas en 1949.[24] Este espécimen se ha ligado al nombre I. mantelli, una especie nombrada en 1832 por von Meyer en lugar de I. anglicus, solamente porque procedía de una formación rocosa diferente que el material original de I. anglicus.[21]
 El famoso banquete de Benjamin Waterhouse Hawkins dentro del Iguanodon.
El famoso banquete de Benjamin Waterhouse Hawkins dentro del Iguanodon.Al mismo tiempo, la tensión comenzó a crecer entre Mantell y Richard Owen, un científico ambicioso con una mayor financiación y mejores conexiones sociales en los turbulentos mundos de la ciencia y la política británicas de la era del Acta de la Reforma de 1832. Owen, un firme creacionista, se oponía al "transmutacionismo", una versión primitiva de la evolución, que entonces estaba siendo discutida y utilizó a los dinosaurios —el nombre lo acuñaría él mismo algo más tarde— como arma en este conflicto. Con la definición de Dinosauria redujo las dimensiones de los dinosaurios desde los excesivos más de 61 metros, y determinó que no eran simplemente lagartos gigantes, proponiendo que eran tan avanzados como los mamíferos, características dadas por Dios, y según la comprensión de su tiempo, no habrían podido ser "transmutados" desde reptiles a criaturas como los mamíferos.[25] [26]
Poco antes su muerte en 1852, Mantell argumentó que Iguanodon no era un pesado animal similar a un paquidermo[27] como Owen proponía, pero no fue invitado a participar en la reconstrucción de los dinosaurios de The Crystal Palace, que impusieron la imagen de los dinosaurios de Owen en la imaginación popular.[25] Junto a Benjamin Waterhouse Hawkins, crearon casi dos docenas de esculturas de tamaño natural de varios animales prehistóricos, construidos en hormigón sobre una estructura de acero y ladrillo. Incluyeron dos Iguanodon, uno erguido y otro a cuatro patas y, antes que este último fuera terminado, se llevó a cabo un banquete en su interior para veinte selectos comensales.[28] [29] [30]
La mina de Bernissart
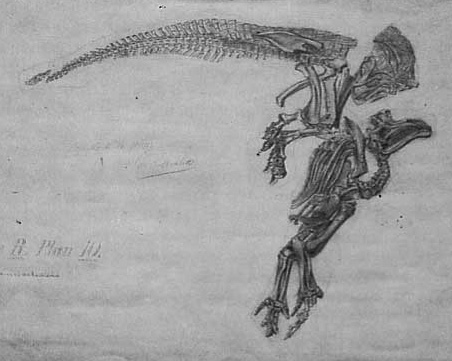
 Fotografía de un esqueleto de Iguanodon de Bernissart siendo montado.
Fotografía de un esqueleto de Iguanodon de Bernissart siendo montado.El mayor descubrimiento de restos de Iguanodon ocurrió en 1878 en la mina de carbón de Bernissart en Bélgica, a una profundidad de 322 metros.[7] Louis Dollo, con Louis de Pauw y el estímulo de Alphonse Briart, supervisor de minas de Morlanwelz, dirigió la excavación y reconstrucción de los esqueletos. Se recuperaron al menos 38 individuos de Iguanodon,[1] la mayoría de ellos adultos.[31] Muchos de ellos fueron preparados y expuestos al público en 1882 y aún pueden verse, en una exhibición impresionante, en el museo del Real Instituto Belga de Ciencias Naturales en Bruselas: 9 ejemplares montados erguidos y otros 19 mostrados como fueron encontrados.[7] Replicas de estos se exhiben en Museo de Historia Natural de la Universidad de Oxford y en el Museo Sedgwick de Ciencias de la Tierra en Cambridge. Otro de los ejemplares originales montados está expuesto en un pequeño museo ad oc en la ciudad de Benissart. La mayoría de los especímenes fueron asignados a una nueva especie, I. bernissartensis, un animal más grande y pesado que los restos encontrados en Inglaterra, pero otro espécimen, más pequeño y grácil, fue asignado a I. mantelli (hoy Dollodon bampingi). Estos restos fueron algunos de los primeros esqueletos completos de dinosaurio conocidos. Junto a los Iguanodon fueron encontrados restos de plantas, insectos, peces, anfibios y de otros reptiles,[7] incluyendo el cocodrilo Bernissartia.[15]
 Iguanodon bernissartensis. Dibujos de los fósiles de 1882.
Iguanodon bernissartensis. Dibujos de los fósiles de 1882.Cuando se dispusieron a reconstruir y conservar los restos de Bernissart, el conocimiento de la conservación de restos fósiles estaba en su infancia, y estaban mal equipados para tratar lo que se conocía como "la enfermedad de la pirita". La pirita (sulfuro de hierro) en los huesos se alteraba a sulfato de hierro dañando los restos, haciéndolos agrietarse y desmenuzándolos. Cuando estaban aislados en la roca, los huesos se encontraban en un ambiente que evitaba el proceso, pero cuando fueron expuestos al aire, la reacción química natural comenzó a ocurrir. Sin conocer la causa verdadera, el personal del museo en Bruselas supuso que se encontraban ante una infección, y trataron el problema con una combinación de alcohol, arsénico, y laca, pensada para penetrar (alcohol), matar cualquier agente biológico (arsénico) y endurecer (laca) simultáneamente los fósiles. Este tratamiento tuvo el efecto involuntario de aislar los restos húmedos y prolongar el período de daño. Los tratamientos modernos en lugar de esto contemplan la eliminación de la humedad de los restos, preparando un baño especial en polietilenglicol, que aplica en caliente dentro de una bomba de vacío, así el polietilenglicol se infiltra en los poros sustituyendo al agua, sellando y consolidando el fósil.[7]
 Uno de los varios esqueletos mostrados en Bruselas.
Uno de los varios esqueletos mostrados en Bruselas.Los especímenes de Dollo permitieron que se demostrara que los paquidermos prehistóricos de Owen no eran la imagen correcta de Iguanodon. En su lugar diseñó los montajes esqueléticos basándose en el emú y el walabí, y puso la púa que había estado en la nariz correctamente en el pulgar.[32] [33] Sus montajes no fue totalmente correctos ya que tenía la desventaja de ser el primero en enfrentarse a un dinosaurio completo. Un problema que fue reconocido más adelante fue la curvatura que introdujo en la cola. Este apéndice era más o menos recto, como se muestra en los esqueletos que él excavaba, y la presencia de tendones osificados. De hecho, para conseguir la curva de la cola más parecida a la del walabí o canguro, tendría que haber estado rota. Con su cola y parte posterior correctas, rectas, el animal habría caminado con su columna vertebral horizontal, paralela a la tierra, con los brazos para apoyar el cuerpo si fuera necesario.[7] Las excavaciones en la mina pararon en 1881, aunque no fuera agotada de fósiles, como han demostrado recientes operaciones de perforación.[34] Durante la Primera Guerra Mundial, cuando la ciudad fue ocupada por los alemanes, se hicieron preparativos para abrir de nuevo la mina para la paleontología y Otto Jaekel fue enviado desde Berlín para supervisarlas. Cuando los aliados recuperaron Bernissart apenas la primera capa fosilífera estaba a punto de ser destapada. Otras tentativas de abrir de nuevo la mina fueron obstaculizadas por problemas financieros y definitivamente paradas en 1921, cuando la mina se inundó.[7]
Descubrimientos en todo el mundo
Las investigaciones sobre Iguanodon disminuyeron durante la primera parte del siglo XX, cuando las Guerra Mundiales y la Gran Depresión envolvieron a Europa.
En 1925 R. W. Hooley describió una nueva especie, I. atherfieldensis, procedente de Atherfield Point en la Isla de Wight, que suscitó una intensa controversia taxonómica.[35]
Sin embargo, el que había sido un género exclusivamente europeo empezó a ser identificado en todo el mundo, con material procedente de África (dientes en Túnez,[36] y en otras partes del Desierto del Sahara),[37] Mongolia (I. orientalis),[38] y en América del Norte (I. ottingeri de Utah, Estados Unidos).[39] Otra especie de norteamérica, de Dakota del Sur, I. lakotaensis,[40] fue posteriormente reasignada al género Dakotadon.[41]
Iguanodon no fue parte del principio del renacimiento de los dinosaurios que comenzó con la descripción del Deinonychus en 1969, pero no fue olvidado por mucho tiempo. El trabajo sobre mecanismos de alimentación de los ornitópodos de David B. Weishampel proporcionó una mejor comprensión de su alimentación,[5] y David B. Norman trabajó muchos aspectos de Iguanodon, haciéndolo uno de los dinosaurios más conocidos.[10] [7] [42] [1] Además, el hallazgo de numerosos esqueletos de Iguanodon, en Nehden, Renania del Norte-Westfalia, Alemania, proveyó la evidencia de comportamiento gregario, en manada, en este género ya que los animales en el área del hallazgo parecían haber estado próximos unos a otros cuando una inundación repentina los mató. Se encontraron al menos 15 individuos, de 2 a 8 metros de largo, aunque algunos de ellos son iguanodóntidos gráciles y pertenecen al género relacionado Mantellisaurus o Dollodon (descrito inicialmente como I. atherfieldensis, por creerse en aquel momento que se trataba de otra especie de Iguanodon).[31] [41]
En 2003, Graham Embery et al.,[43] en búsqueda de ADN y otras biomoleculas conservadas en fósiles, encontraron en una costilla de Iguanodon restos identificables de proteínas típicas del tejido óseo, como fosfoproteínas y proteoglicanos.
Clasificación
Cladística

Iguanodon le da al nombre al clado sin clasificar Iguanodontia, un grupo muy populoso de ornitopódos con muchas especies conocidas del Jurásico Medio al Cretácico Superior. De este grupo, Iguanodon es el mejor conocido, en el cual se incluyen también Dryosaurus, Camptosaurus, Ouranosaurus, y los dinosaurios pico de pato o hadrosáuridos. En fuentes antiguas, Iguanodontidae aparecía como una familia independiente.[44] [45] Esta familia ha sido usada tradicionalmente como un taxón cajón de sastre, incluyendo ornitopódos que no eran hipsilofodóntidos o hadrosáuridos. En la práctica, animales como Callovosaurus, Camptosaurus, Craspedodon, Kangnasaurus, Mochlodon, Muttaburrasaurus, Ouranosaurus y Probactrosaurus fueron usualmente asignados a esta familia.[45] Con el advenimiento del análisis cladistico se demostró que Iguanodontidae, como se estaba interpretado tradicionalmente, era un grupo parafilético y que en un cladograma, refiriéndolos a los hadrosáuridos, estos animales se separaban en diversos clados, en vez de estar agrupados en un único clado.[12] [1] Esencialmente, el concepto moderno de Iguanodontidae incluye solamente al género Iguanodon. Los grupos como Iguanodontoidea todavía se utilizan como clados sin clasificar en la literatura científica, aunque muchos iguanodóntidos tradicionales ahora se incluyen en la superfamilia Hadrosauroidea. Iguanodon es ubicado entre Camptosaurus y Ouranosaurus en cladogramas, y desciende probablemente de un animal similar al camptosaurio. [1] En este punto, Jack Horner sugiere, basado sobre todo en las características del cráneo, que los hadrosáuridos forman dos grupos relacionados separados, con Iguanodon en la línea de los de las cabezas aplanadas hadrosaurinos, y Ouranosaurus en la línea de los crestados lambeosaurinos, [46] pero esta propuesta ha sido rechazada.[12] [1]
Especies
 Iguanodon bernissartensis montado en su postura cuadrúpeda correcta, museo del Real Instituto Belga de Ciencias Naturales, Bruselas.
Iguanodon bernissartensis montado en su postura cuadrúpeda correcta, museo del Real Instituto Belga de Ciencias Naturales, Bruselas.Debido a que Iguanodon es uno de los primeros géneros de dinosaurio en haber recibido un nombre, son numerosas las especies que le han sido asignadas. Si bien Iguanodon no se considera un taxón cajón de sastre (al estilo de otros géneros de dinosaurio como Megalosaurus o Pelorosaurus) su historia ha sido compleja y su taxonomía es revisada constantemente.[47] [48] [49] [41] Los restos de las especies más conocidas han surgido en Bélgica, Inglaterra, España y Francia. Restos de animales similares, posiblemente pertenecientes a este género han sido encontrados también en Túnez y Mongolia, y de especies diferenciadas en Utah, Estados Unidos. Gregory Paul ha recomendado limitar el uso de I. bernissartensis estrictamente a los descubrimientos de Bernissart, y utilizar Iguanodon sp. (que significa especie indeterminada) para los restos de robustos animales tipo iguanodonte de otros sedimentos europeos del Barremiano.[41] Al final, lo que inicialmente se pensó que era un dinosaurio típico de las islas Británicas puede en realidad haber sido escaso.
Iguanodon anglicus fue la especie tipo original, pero el holotipo estaba basado en un solo diente y desde entonces sólo se han encontrado restos parciales de la especie. En marzo de 2000, la Comisión Internacional de Nomenclatura Zoológica cambió la especie tipo a la mejor conocida I. bernissartensis. El diente del iguanodonte original es guardado en el Te Papa Tongarewa, el museo nacional de Nueva Zelanda en Wellington, aunque no está exhibida actualmente. El fósil llegó a Nueva Zelanda luego que el hijo de Gideon Mantell se estableciera allí.[50]
Especies reconocidas de Iguanodon
Sólo unas pocas de las especies originalmente asignadas a la familia de los iguanodóntidos son actualmente consideradas válidas, y de ellas, sólo una corresponde al género Iguanodon.[1] [41] I. bernissartensis, fue descrito por George Albert Boulenger en 1881, siendo el neotipo para el género. Esta especie es la más conocida por los numerosos esqueletos completos descubiertos en Bernissart, pero también se conoce por abundantes restos de otras localidades de Europa.
David Norman sugiere que se incluya a la dudosa I. orientalis de Mongolia,[51] pero esto no ha sido refrendado por otros investigadores.[41]
Especies reasignadas
Dos especies de iguanodonte inicialmente propuestas por Richard Owen como Iguanodon han sido reasignadas a otros géneros. La primera, I. hoggi, bautizada por Owen en base a una mandíbula inferior hallada en sedimentos del Grupo Purbeck del Titoniano-Berriasiano (Jurásico superior-Cretácico inferior) en Dorset en 1874, fue finalmete asignada al género Owenodon[52] (previamente había sido atribuida a Camptosaurus por David Norman y Paul Barrett).[48] La segunda, I. major, basada en una vértebra encontrada en la Isla de Wight fue descrita por Owen en 1842 como una especie de Streptospondylus, es todavía un nomen dubium, pero podría ser un sinónimo de I. anglicus,[1] u otra especie independiente.[53]
Adicionalmente, otras dieciséis especies consideradas originalmente como iguanodontes han sido reclasificadas en otros géneros. La especie Albisaurus albinus, un arcosaurio no dinosauriano, fue descrita inicalmente en 1893, basándose en escasos restos, por el paleontólogo checo Antonin Fritsch (Frič en su ortografía original) como Iguanodon albinus.[54]
I. atherfieldensis, descrita por Reginald Hooley en 1925,[35] era más pequeño y menos robusto que I. bernissartensis, con una columna vertebral más larga, pero fue renombrada como Mantellisaurus atherfieldensis en 2007.[49]
I. exogyrarum fue descrita por Fritsch en 1878. Es un nomen dubium en base al poco material que existe de ella y fue reasignada por George Olshevsky, como Ponerosteus.[55]
I. valdensis, descrita por Hulke en 1879 a partir de restos de vértebras y de la pelvis, encontrados en la Isla de Wight en una capa del Barremiano,[56] originalmente llamada Vectisaurus, puede ser un espécimen juvenil de Mantellisaurus atherfieldensis,[57] o de una especie no determinada de Mantellisaurus.[41]
I. gracilis, bautizada por Lydekker en 1888 como especie tipo de Sphenospondylus y asignada como Iguanodon en 1969 por Rodney Steel, podría también pertenecer a Mantellisaurus atherfieldensis.[1]
Otras dos especies descritas por Richard Lydekker al final del siglo XIX han sido reasignadas a otros géneros.[58] I. dawsoni, descrita por Lydekker en 1888,[59] y conocida por dos esqueletos parciales encontrados en la Formación Arcillas de Wadhurst (Sussex del Este, Inglaterra),[1] de mediados del Valanginiano (Cretácico inferior),[41] es actualmente la especie tipo del género Barilium.[58] I. fittoni, procedente de la misma formación,[1] descrita por Lydekker in 1889,[60] es ahora la especie tipo de Hypselospinus.[58]
I. foxii fue descrita originalmente por Thomas Henry Huxley en 1869 como especie tipo de Hypsilophodon; Owen (1873 o 1874) la reasignó a Iguanodon, pero su asignación fue rápidamente contestada.[61]
I. hollingtoniensis, descrita por Lydekker en 1889, ha sido considerada como sinónimo de I. fittoni.[1] Otro espécimen asignado por Owen a I. hollingtonensis en 1874, con una combinación inusual de mandíbula inferior tipo hadrosaúrido y un torax robusto, puede representar un taxón no clasificado.[41]
I. prestwichii, desrito por John Hulke en 1880, fue reasignado a Camptosaurus prestwichii.
I. seeleyi, descrito por Hulke dos años después de I. prestwichii, has sido considerado sinónimo de I. bernissartensis,[1] aunque esto todavía sigue en disputa.[41] [53]
I. suessii, descrito por Emanuel Bunzel en 1871, fue reasignado como Mochlodon suessi.[1]
I. lakotaensis, que fue descrito por David B. Weishampel y Philip R. Bjork en 1989,[40] era la única especie de iguanodonte de América del Norte ampliamente aceptada. Fue descrita a partir de un cráneo parcial encontrado en la Formación Lakota de Dakota del Sur (EE.UU.), de edad Barremiano (Cretácico inferior). Su asignación ha generado controversia, pues algunos autores sugieren que es más basal que I. bernissartensis, y lo relacionan con el género Theiophytalia.[62] pero David Norman ha sugerido que puede ser un sinónimo de I. bernissartensis.[47] Posteriormente, Gregory S. Paul le asignó su propio género, Dakotadon.[41]
 IRSNB 1551, el holotipo de Dollodon bampingi, anteriormente Iguanodon mantelli, mostrado en el Real Instituto Belga de Ciencias Naturales en Bruselas.
IRSNB 1551, el holotipo de Dollodon bampingi, anteriormente Iguanodon mantelli, mostrado en el Real Instituto Belga de Ciencias Naturales en Bruselas.Iguanodon mantelli, descrito por von Meyer en 1832, está basado en el mismo material que I. anglicus.[63] Sin embargo, algunos esqueletos, incluyendo el espécimen de Maidstone y uno de los de Bernissart han sido durante muchos años asignados a esta especie, pero su atribución no ha sido definitiva. El esqueleto grácil Bernissart, por ejemplo, fue reasignado, primero como Mantellisaurus atherfieldensis,[42] y, luego de una revisión más exhaustiva, a su propio género y especie, Dollodon bampingi.[41]
I. orientalis, descrito por Rozhdestvensky en 1952,[38] se basó en muy poco material, pero un cráneo y un hocico arqueado que le estaban asignados fueron reclasificados como pertenecientes a Altirhinus kurzanovi en 1998.[47] Al mismo tiempo, I. orientalis se consideró como un nomen dubium no distinguible de I. bernissartensis.[51] [47]
Harry Seeley describió I. phillipsi en 1869,[64] pero posteriormente lo reasignó a Priodontognathus.[65]
En 2011, se identificó un nuevo iguanodontiano del Cretácico de España, Delapparentia turolensis, basándose en un espécimen que fue previamente asignado a Iguanodon bernissartensis[66]
Especies dudosas
Cinco especies de iguanodonte son consideradas como nomina dubia o no descritas. I. anglicus, descrita por Friedrich Holl en 1829,[22] es la especie tipo original de Iguanodon, pero, como se mencionó antes, fue remplazada por I. bernissartensis, que es ahora el neotipo. En el pasado ha sido escrita como I. angelicus (Lessem y Glut, 1993) e I. anglicum (Holl, 1829 corregido, Bronn, 1850). Los únicos restos son los dientes encontrados en el bosque de Tilgate de Sussex del Este, Inglaterra del Valanginiano-Barremiano, del Cretácico inferior. I. hillii, bautizada por Edwin Tully Newton en 1892 en base a un diente encontrado en la Caliza inferior de Hertfordshire, del Cenomaniano (Cretácico superior), es un hadrosáuridos.[67] Los restos de "I. mongolensis" (Whitfield, 1992, nomen nudum de un comentario en la foto de un libro) fueron posteriormente nombrados Altirhinus.[68]
I. ottingeri, descrito por Peter Galton y James A. Jensen en 1979, es un nomen dubium basado en dientes encontrados en la porción infeior de la Formación Cedar Mountain de Utah, probablemente del Aptiano.[39] E. Sauvage en 1876 identificó algunos dientes procedentes de rocas del Jurásico superior de Pas-de-Calais (Francia) como I. praecursor, pero actualmente se interpretan como de saurópodo, incluso han sido identificados como de Neosodon,[69] a pesar de que proceden de formaciones diferentes.[70]
Finalmente, otros géneros y especies mal conocidos también se incluyen en Iguanodon sin ser especies separadas, aunque su asignación podría cuestionarse con la reasignación de I. atherfieldensis. Estos incluyen Heterosaurus neocomiensis (Cornuel, 1850), Hikanodon (Keferstein, 1834), y Therosaurus (Fitzinger, 1840), y las especies Streptospondylus recentior (Owen, 1851), Cetiosaurus brachyurus,[2] y parte de C. brevis (Owen, 1842; C. brevis es una quimera paleontológica).[71] El nomen nudum Proiguanodon (van den Broeck, 1900) también está en este grupo,[72] y posiblemente también los poco conocidos Streptospondylus grandis (Owen, 1851) y S. meyeri (Owen, 1854).[21]
Paleobiología
Alimentación y dieta
Se desconoce exactamente de que se alimentaban los iguanodontes con sus robustas mandíbulas. Algunos autores, como David Norman, han sugerido que su dieta podría estar basada en equisetos, cicas y coníferas;[7] el tamaño de I. bernissartensis le habría permitido alcanzar el follaje de árboles de hasta cuatro o cinco metros de altura.[3] Según algunos autores, los iguanodóntidos en general habrían contribuído al desarrollo de las angiospermas (plantas con flores) durante el Cretácico, pues debido a un extensivo ramoneo de gimnospermas (la flora dominante) habrían proporcionado espacio para el crecimiento de las angiospermas primitivas,[73] aunque no todos los especialistas aceptan esta hipótesis.[74] [1]
Los iguanodontes, debido a su tamaño y abundancia, están considerados como el herbívoro de talla media a grande dominante en su paleobiocenosis.[1] En Inglaterra, por ejemplo, ésta paleocomunidad estaba compuesta, además de Iguanodon, por el pequeño depredador Aristosuchus, los grandes depredadores Eotyrannus, Baryonyx y Neovenator, herbívoros pequeños como Hypsilophodon y Valdosaurus, el también iguanodóntido Mantellisaurus, el dinosaurio acorazado Polacanthus, y saurópodos como Pelorosaurus.[75]
Postura y locomoción
 Recreación de Iguanodon por Samuel Griswold Goodrich para Illustrated Natural History of the Animal Kingdom (1859).[76]
Recreación de Iguanodon por Samuel Griswold Goodrich para Illustrated Natural History of the Animal Kingdom (1859).[76] Pintura de 1905 mostrando a Iguanodon en una pose «trípode» actualmente descartada (Joseph Smit, From nebula to man).
Pintura de 1905 mostrando a Iguanodon en una pose «trípode» actualmente descartada (Joseph Smit, From nebula to man).Estos ornitópodos podían adoptar una postura bípeda, acercando los brazos al suelo, ya que la columna vertebral se equilibraba horizontalmente, así podían caminar en dos o cuatro patas. La cola también estaba en posición horizontal, estirada y bastante rígida por encima del suelo. En un comienzo, los científicos pensaron que los iguanodontes eran animales torpes, como un rinoceronte, al caminar en cuatro patas. Más tarde, cuando en Bélgica se encontraron ciertos esqueletos, creyeron erróneamente que se desplazaban erguidos con la cola apoyada en el suelo. Sus brazos eran largos y sus dedos se doblaban hacía atrás para sostener el cuerpo al caminar en cuatro patas. El quinto dedo era delgado y flexible.
En su revisión del género, David Norman demostró que la clásica postura vertical de Iguanodon arrastrando la cola a modo de tercera pierna, según habían interpretado Dollo y sus colaboradores a partir de los fósiles de Bernissart, era una postura imposible debido a la presencia de los tendones osificados a lo largo buena parte de la columna vertebral:[10] para adquirir la postura en trípode, la cola debería literalmente romperse.[7] Al presentar al animal con la columa vertebral recta y horizontal consiguió que muchos aspectos de los robustos brazos y cintura escapular fueran más comprensibles. Por ejemplo, la mano y la muñeca eran relativamente rígidas y capaces de la hiperextensión, con las falanges de los tres dedos centrales agrupadas formando una especie de pezuña, le habrían permitido soportar su peso repartido entre las cuatro extremidades. Todas estas características sugieren que la postura habitual de estos animales fuera cuadrúpeda.[10]
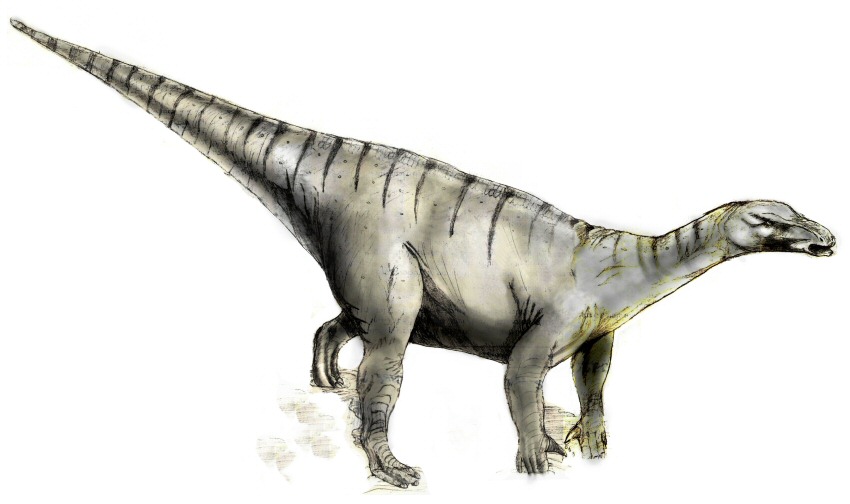
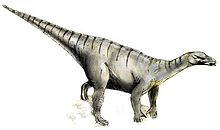 Ilustración de Iguanodon como cuadrúpedo.
Ilustración de Iguanodon como cuadrúpedo.Por otra parte, parece que la preferencia por posición cuadrúpeda de los iguanodontes se incrementaba con la edad, según se hacían más pesados; durante la etapa juvenil, I. bernissartensis, tenía los brazos proporcionalmente más cortos que en la etapa adulta (el 60% de la longitud de las patas contra el 70% en los adultos).[1] Los pies de tres dedos de los iguanodontos eran relativamente largos.
Al caminar como cuadrúpedo, el animal habría llevado las manos con las palmas enfrentadas, como se muestra por las icnitas y la anatomía de brazos y manos,[77] [78] y habría apoyado únicamente los dedos, tanto los de las extremidades delanteras como los de las traseras (a la manera digitígrada).[1] La velocidad máxima estimada para los iguanodontes es de 6,7 m/s (unos 24 km/h)[79] en modo bípedo, pues como cuadrúpedo no habría podido galopar.[1] Un rastro de icnitas en Inglaterra muestra lo que pudiera ser la marcha tetrápoda de un iguanodonte, aunque las impresiones de los pies son pobres, siendo su atribución directa incierta.[10]
En sedimentos cretácicos de toda Europa se han encontrado huellas fósiles (icnitas) atribuidas a Iguanodon, en muchos casos en regiones en las que también se encuentran huesos fosilizados del este género.,[80] [81] sin embargo la atribución de unas huellas a un taxón determinado tiene un cierto grado de incertidumbre y no hay pruebas directas que permitan atribuir estas huellas a Iguanodon.[7] Desde antiguo se conocen huellas tridáctilas en rocas sedimentarias del Cretácico Inferior de Gran Bretaña, especialmente en capas del Grupo Wealden en la Isla de Wight. Aunque difíciles de interpretar en un principio, algunos autores las asociaron con dinosaurios, y en 1846, E. Tagert fue más lejos, asignándolas a un icnogénero que denominó directamente como Iguanodon.[82] Un poco más tarde, Samuel Beckles, publicó en 1854 que aunque tenían el aspecto de pisadas de ave, bien pudieran haber sido originadas por dinosaurios.[83] La posible identidad de los autores de estas huellas quedó mucho más evidente tras el descubrimiento, en 1857, de los huesos de una pata de un Iguanodon juvenil, cuyo pie mostraba claramente tres únicos dedos y que posibilitaban, por tanto, aquellas huellas tridáctilas.[84] [85]
Comportamiento social
Parece que Iguanodon bernissartensis pudo haber tenido hábitos gregarios, pues los hallazgos de Nehden muestran un amplio abanico de edades individuales, a lo que se suma la presencia de especies similares como Dollodon o Mantellisaurus. La naturaleza geográfica confinada pudo haber registrado la mortalidad de una manada de animales que emigraban a través de ríos.[31]
De igual manera se había interpretado la acumulación de restos de Bernissart, como resultado de una sola catástrofe, si bien ahora se interpreta como resultado de múltiples eventos. Según los nuevos análisis, se registran por lo menos tres sucesos de alta mortalidad y, aunque numerosos individuos habrían muerto en un periodo geológicamente breve (10-100 años),[31] no prueba necesariamente que estos iguanodontes vivieran en manadas.[1] Otro elemento contra los indicios de gregarismo en Bernissart es que los restos juveniles son muy infrecuentes, a diferencia de casos modernos de mortalidad en masa de una manada. Eran más probablemente víctimas de inundaciones repentinas periódicas, cuyos cadáveres se acumulaban en una zona lacustre o pantanosa.[31]
A diferencia de otros herbívoros gregarios no hay evidencias que apoyen un marcado dimorfismo sexual, como en hadrosáuridos o ceratopsios. En un tiempo se llegó a sugerir que I. mantelli o I. atherfieldensis (actualmente Dollodon y Mantellisaurus, respectivamente) de Bernissart representaban hembras y los más grandes y robustos I. bernissartensis machos de una misma especie,[86] pero se descartó posteriormente.[10] [7] [49]
En la cultura popular
 Estatua de Iguanodon por Benjamin Waterhouse Hawkins, todavía visible en The Crystal Palace, y un modelo moderno de Iguanodon del Museo de Historia Natural de la Universidad de Oxford.
Estatua de Iguanodon por Benjamin Waterhouse Hawkins, todavía visible en The Crystal Palace, y un modelo moderno de Iguanodon del Museo de Historia Natural de la Universidad de Oxford.Desde su descripción en 1825, Iguanodon fue mundialmente conocido y las dos estatuas de tamaño natural que se construyeron para el Palacio de Cristal en Londres en 1852 contribuyeron en gran medida a su popularidad.[87] Con las falanges de los pulgares como cuernos en la nariz, sobre cuatro patas como un elefante, fueron la primera reconstrucción en tamaño natural de un dinosaurio.
 Estatua de Iguanodon en la entrada al acuario del Zoológico de Berlín.
Estatua de Iguanodon en la entrada al acuario del Zoológico de Berlín.Ya que fue uno de los primeros dinosaurios descritos y es uno de los dinosaurios más conocidos, Iguanodon ha servido como «barómetro» del cambio de la percepción científica y popular sobre los dinosaurios. Sus reconstrucciones han pasado a través de tres etapas: el reptil-elefante cuadrúpedo y con un cuerno sobre el hocico de la era victoriana; un bípedo, pero todavía lento y torpe, que usa su cola para apoyarse y, finalmente, su representación más ágil y dinámica, en la que se desplaza tanto a dos como a cuatro patas. La segunda representación dominó el siglo XX, pero fue cambiando a partir de los años 70.[88]
Han jugado un papel estelar en el libro de Arthur Conan Doyle El mundo perdido (1912). También apareció en Red Raptor (1995) de Robert Bakker, como presa de un Utahraptor.
Varias películas han representado iguanodontes. En la película de Disney Dinosaurio, un iguanodonte llamado Aladar sirvió de protagonista junto a otros tres iguanodontes y muchos otros personajes. Junto a Tyrannosaurus y Stegosaurus es uno de los tres dinosaurios que inspiraron a Godzilla.[89] Los iguanodontes también aparecen en la serie de películas de En Busca del Valle Encantado y en algunos episodios de la serie homónima para televisión.
Aparte de sus apariciones en el mundo del cine, Iguanodon también ha aparecido en televisión, como en la miniserie documental Caminando con dinosaurios (1999) producida por la BBC, donde es mostrado que vivían por todo el mundo, eran cazados por utarraptores y vivían asociados a dinosaurios nodosáuridos como Polacanthus.
Entre otros dinosaurios, se recrea un iguanodonte en una de las atracciones del parque temático Animal Kingdom (Walt Disney World, Florida, Estados Unidos).
En honor al género se denominó (9941) Iguanodon a un asteroide del cinturón de asteroides principal.[90] [91]
Véase también
Referencias
- ↑ a b c d e f g h i j k l m n ñ o p q r s t u v w x y z aa Norman, David B. (2004). «Basal Iguanodontia». En Weishampel, D.B., Dodson, P., and Osmólska, H. (eds.). The Dinosauria (2nd edición). Berkeley: University of California Press. pp. 413–437. ISBN 0-520-24209-2.
- ↑ a b c Glut, Donald F. (1997). «Iguanodon». Dinosaurs: The Encyclopedia.. Jefferson, North Carolina: McFarland & Co. pp. 490–500. ISBN 0-89950-917-7.
- ↑ a b c d e f Naish, Darren; David M. Martill (2001). «Ornithopod dinosaurs». Dinosaurs of the Isle of Wight. London: The Palaeontological Association. pp. 60–132. ISBN 0-901702-72-2.
- ↑ Blows, W. T. (1997). «A review of Lower and middle Cretaceous dinosaurs from England». En Lucas, S.G., Kirkland, J.I., and Estep, J.W. (eds.). Lower and Middle Cretaceous Terrestrial Ecosystems. New Mexico Museum of Natural History and Science Bulletin, 14. New Mexico Museum of Natural History and Science. pp. 29–38.
- ↑ a b Weishampel, David B. (1984). Evolution in jaw mechanics in ornithopod dinosaurs. Advances in Anatomy, Embryology, and Cell Biology, 87. Berlin; New York: Springer-Verlag. ISBN 0387131140.
- ↑ Bakker, Robert T. (1986). «Dinosaurs At Table». The Dinosaur Heresies. New York: William Morrow. pp. 160–178. ISBN 0140100555.
- ↑ a b c d e f g h i j k l m n Norman, David B. (1985). «To Study a Dinosaur». The Illustrated Encyclopedia of Dinosaurs: An Original and Compelling Insight into Life in the Dinosaur Kingdom. New York: Crescent Books. pp. 24–33. ISBN 0-517-468905.
- ↑ a b c Mantell, Gideon A. (1825). «Notice on the Iguanodon, a newly discovered fossil reptile, from the sandstone of Tilgate forest, in Sussex.». Philosophical Transactions of the Royal Society 115: pp. 179–186. doi:. http://links.jstor.org/sici?sici=0261-0523(1825)115%3C179%3ANOTIAN%3E2.0.CO%3B2-W.
- ↑ Mantell, Gideon A. (1848). «On the structure of the jaws and teeth of the Iguanodon». Philosophical Transactions of the Royal Society of London 138: pp. 183–202. doi:. http://links.jstor.org/sici?sici=0365-0855(1843%2F1850)5%3C757%3AOTSOTJ%3E2.0.CO%3B2-K.
- ↑ a b c d e f Norman, David B. (1980). «On the ornithischian dinosaur Iguanodon bernissartensis of Bernissart (Belgium)». Mémoires de l'Institut Royal des Sciences Naturelles de Belgique 178: pp. 1–105.
- ↑ Norman, D.B. (1985). The Illustrated Encyclopedia of Dinosaurs, 115.
- ↑ a b c Norman, David B.; Weishampel, David B. (1990). «Iguanodontidae and related ornithopods». En Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.).. The Dinosauria. Berkeley: University of California Press. pp. 510–533. ISBN 0-520-06727-4.
- ↑ Galton, Peter M. (1973). «The cheeks of ornithischian dinosaurs». Lethaia 6: pp. 67–89. doi:.
- ↑ Fastovsky, D.E., and Smith, J.B. "Dinosaur paleoecology." The Dinosauria, 614–626.
- ↑ a b Bultynck, Pierre (1989) (en francés). Bernissart et les Iguanodons. Lovaina: Institut Royal des Sciences Naturelles de Belgique. pp. 115.
- ↑ Tweedie, Michael W.F. (1977). The World of the Dinosaurs. London: Weidenfeld & Nicolson. pp. 143. ISBN 0688032222.
- ↑ Naish, D. «Venomous & Septic Bites». Consultado el 14-02-2007.
- ↑ a b Sues, Hans-Dieter (1997). «European Dinosaur Hunters». En James Orville Farlow and M. K. Brett-Surman (eds.). The Complete Dinosaur. Bloomington: Indiana University Press. pp. 14. ISBN 0-253-33349-0.
- ↑ Lucas, Spencer G. (December 1999). «Book review: Gideon Mantell and the discovery of dinosaurs». Palaios 14 (6): pp. 601–602. doi:. ISSN 0883-1351. http://links.jstor.org/sici?sici=0883-1351%28199912%2914%3A6%3C601%3AGMATDO%3E2.0.CO%3B2-I.
- ↑ Cadbury, D. (2000). The Dinosaur Hunters. Fourth Estate:London, 384 p. ISBN 1-85702-959-3.
- ↑ a b c Olshevsky, G. «Re: Hello and a question about Iguanodon mantelli (long)». Consultado el 11-02-2007.
- ↑ a b Holl, Friedrich (1829). Handbuch der Petrifaktenkunde, Vol. I. Ouedlinberg. Dresden: P.G. Hilscher. OCLC 7188887.
- ↑ Mantell, Gideon A. (1834). «Discovery of the bones of the Iguanodon in a quarry of Kentish Rag (a limestone belonging to the Lower Greensand Formation) near Maidstone, Kent.». Edinburgh New Philosophical Journal 17: pp. 200–201.
- ↑ Colbert, Edwin H. (1968). Men and Dinosaurs: The Search in Field and Laboratory. New York: Dutton & Company. ISBN 0140212884.
- ↑ a b Torrens, Hugh. "Politics and Paleontology". The Complete Dinosaur, 175–190.
- ↑ Owen, R. (1842). «Report on British Fossil Reptiles: Part II.». Report of the British Association for the Advancement of Science for 1841 1842: pp. 60–204.
- ↑ Mantell, Gideon A. (1851). Petrifications and their teachings: or, a handbook to the gallery of organic remains of the British Museum.. London: H. G. Bohn. OCLC 8415138.
- ↑ Benton, Michael S. (2000). «brief history of dinosaur paleontology». En Gregory S. Paul (ed.). The Scientific American Book of Dinosaurs. New York: St. Martin's Press. pp. 10–44. ISBN 0-312-26226-4.
- ↑ Yanni, Carla (September 1996). «Divine Display or Secular Science: Defining Nature at the Natural History Museum in London». The Journal of the Society of Architectural Historians 55 (3): pp. 276–299. doi:.
- ↑ Norman, David B. The Illustrated Encyclopedia of Dinosaurs. p. 11.
- ↑ a b c d e Norman, David B. (marzo 1987). «A mass-accumulation of vertebrates from the Lower Cretaceous of Nehden (Sauerland), West Germany». Proceedings of the Royal Society of London. Series B, Biological Sciences 230 (1259): pp. 215–255. doi:.
- ↑ Dollo, Louis (1882). «Première note sur les dinosauriens de Bernissart». Bulletin du Musée Royal d'Histoire Naturelle de Belgique 1: pp. 161–180 idioma=francés.
- ↑ Dollo, Louis (1883). «Note sur les restes de dinosauriens recontrés dans le Crétacé Supérieur de la Belgique». Bulletin du Musée Royal d'Histoire Naturelle de Belgique 2: pp. 205–221 idioma=francés.
- ↑ de Ricqlès, A. (2003). «Bernissart's Iguanodon: the case for "fresh" versus "old" dinosaur bone.». Journal of Vertebrate Paleontology 23 (Supplement to Number 3): pp. 45A. ISSN 0272-4634. Abstracts of Papers, Sixty-Third Annual Meeting.
- ↑ a b Hooley, R. W. (1925). «On the skeleton of Iguanodon atherfieldensis sp. nov., from the Wealden Shales of Atherfield (Isle of Wight)». Quarterly Journal of the Geological Society of London 81 (2): pp. 1–61. ISSN 0370-291X.
- ↑ Lapparent, A. F. de (1951). «Découverte de Dinosauriens associés à une faune de Reptiles et de Poissons, dans le Crétacé inférieur de l'Extrême Sud Tunisien» (en francés). Comptes Rendus Hebdomadaires des Séances de l'Académie des Sciences 232: pp. 1430. ISSN 0567-655X.
- ↑ Lapparent, A. F. de (1960). «Les dinosauriens du "Continental Intercalaire" du Sahara Central» (en francés). Mémoires de la Socété Géologique de France (Nouvelle Série) 88A: pp. 1–57.
- ↑ a b Rozhdestvensky, A.K. (1952). «[Discovery of an iguanodont in Mongolia]» (en ruso). Dokladi Akademii Nauk S.S.S.R. 84 (6): pp. 1243–1246.
- ↑ a b Galton, P. M.; J. A. Jensen (1979). «Remains of ornithopod dinosaurs from the Lower Cretaceous of North America». Brigham Young University Geology Studies 25 (3): pp. 1–10. ISSN 1041-7184.
- ↑ a b Weishampel, David B.; Phillip R. Bjork (1989). «The first indisputable remains of Iguanodon (Ornithischia: Ornithopoda) from North America: Iguanodon lakotaensis, sp. nov.». Journal of Vertebrate Paleontology 9 (1): pp. 56–66.
- ↑ a b c d e f g h i j k l Paul, Gregory S. (2008). «A revised taxonomy of the iguanodont dinosaur genera and species». Cretaceous Research 29 (2): pp. 192–216. doi:.
- ↑ a b Norman, David B. (1986). «On the anatomy of Iguanodon atherfieldensis (Ornithischia: Ornithopoda)». Bulletin de L'institut Royal des Sciences Naturelles de Belgique Sciences de la Terre 56: pp. 281–372. ISSN 0374-6291.
- ↑ Embery, Graham; Milner, Angela C.; Waddington, Rachel J.; Hall, Rachel C.; Langley, Martin S.; and Milan, Anna M. (2003). «Identification of proteinaceous material in the bone of the dinosaur Iguanodon». Connective Tissue Research (Suppl. 1): pp. 41–46. doi:. PMID 12952172.
- ↑ Galton, Peter M. (September 1974). «Notes on Thescelosaurus, a conservative ornithopod dinosaur from the Upper Cretaceous of North America, with comments on ornithopod classification». Journal of Paleontology 48 (5): pp. 1048–1067. http://links.jstor.org/sici?sici=0022-3360%28197409%2948%3A5%3C1048%3ANOTACO%3E2.0.CO%3B2-L.
- ↑ a b Norman, David B. "Iguanodontidae". The Illustrated Encyclopedia of Dinosaurs, 110–115.
- ↑ Horner, J. R. (1990). «Evidence of diphyletic origination of the hadrosaurian (Reptilia: Ornithischia) dinosaurs». En Kenneth Carpenter and Phillip J. Currie (eds.). Dinosaur Systematics: Perspectives and Approaches. Cambridge: Cambridge University Press. pp. 179–187. ISBN 0-521-36672-0.
- ↑ a b c d Norman, David B. (January 1998). «On Asian ornithopods (Dinosauria, Ornithischia). 3. A new species of iguanodontid dinosaur». Zoological Journal of the Linnean Society 122 (1–2): pp. 291–348. doi:.
- ↑ a b Norman, David B.; Barrett, Paul M. (2002). «Ornithischian dinosaurs from the Lower Cretaceous (Berriasian) of England». En Milner, Andrew, and Batten, David J. (eds.). Life and Environments in Purbeck Times. Special Papers in Palaeontology 68. London: Palaeontological Association. pp. 161–189. ISBN 0901702730.
- ↑ a b c Paul, Gregory S. (2007). «Turning the old into the new: a separate genus for the gracile iguanodont from the Wealden of England». En Kenneth Carpenter (ed.). Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs. Bloomington: Indiana University Press. pp. 69–77. ISBN 0-253-34817-X.
- ↑ Royal Society of New Zealand. Celebrating the great fossil hunters. Archivado del original el 2005-08-26. http://web.archive.org/web/20050826130408/http://www.rsnz.org/topics/biol/dna50/breakfast.php. Consultado el 22-02-2007.
- ↑ a b Norman, David B. (March 1996). «On Asian ornithopods (Dinosauria, Ornithischia). 1. Iguanodon orientalis Rozhdestvensky, 1952». Zoological Journal of the Linnean Society 116 (2): pp. 303–315. doi:.
- ↑ «Notes on Neocomian (Late Cretaceous) ornithopod dinosaurs from England - Hypsilophodon, Valdosaurus, "Camptosaurus", "Iguanodon" – and referred specimens from Romania and elsewhere». Revue de Paléobiologie 28 (1): 211–273. 2009.
- ↑ a b Naish, Darren; and Martill, David M. (2008). «Dinosaurs of Great Britain and the role of the Geological Society of London in their discovery: Ornithischia». Journal of the Geological Society, London 165: pp. 613–623.
- ↑ Brikman, Winand (1988) (en alemán). Zur Fundgeschichte und Systematik der Ornithopoden (Ornithischia, Reptilia) aus der Ober-Kreide von Europa. Documenta Naturae, 45. Munich: Kanzler. ISBN 3865440452.
- ↑ Olshevsky, George (2000). An annotated checklist of dinosaur species by continent. Mesozoic Meanderings, 3. San Diego: G. Olshevsky Publications Requiring Research. OCLC 44433611.
- ↑ Galton, P.M. (1976). «The Dinosaur Vectisaurus valdensis (Ornithischia: Iguanodontidae) from the Lower Cretaceous of England». Journal of Paleontology 50 (5): pp. 976–984. http://links.jstor.org/sici?sici=0022-3360%28197609%2950%3A5%3C976%3ATDVV%28I%3E2.0.CO%3B2-H.
- ↑ Norman, David B. "A review of Vectisaurus valdensis, with comments on the family Iguanodontidae". Dinosaur Systematics, 147–161.
- ↑ a b c Norman, David B. (2010). «A taxonomy of iguanodontians (Dinosauria: Ornithopoda) from the lower Wealden Group (Cretaceous: Valanginian) of southern England». Zootaxa 2489: pp. 47–66. http://mapress.com/zootaxa/2010/f/z02489p066f.pdf.
- ↑ Lydekker, Richard (1888). «Note on a new Wealden iguanodont and other dinosaurs». Quarterly Journal of the Geological Society of London 44: 46–61. doi:.
- ↑ Lydekker, Richard (1889). «On the remains and affinities of five genera of Mesozoic reptiles». Quarterly Journal of the Geological Society of London 45: 41–59. doi:.
- ↑ Woodward, Henry (1885). «On Iguanodon mantelli, Meyer». Geological Magazine, series 3 2 (1): pp. 10–15. OCLC 2139602.
- ↑ Brill, Kathleen and Kenneth Carpenter. "A description of a new ornithopod from the Lytle Member of the Purgatoire Formation (Lower Cretaceous) and a reassessment of the skull of Camptosaurus." Horns and Beaks, 49–67.
- ↑ Olshevsky, G. «Re: Hello and a question about Iguanodon mantelli (long)». Consultado el 11-02-2007.
- ↑ Seeley, Harry G. (1869). Index to the fossil remains of Aves, Ornithosauria, and Reptilia, from the secondary system of strata arranged in the Woodwardian Museum of the University of Cambridge. Cambridge: Deighton, Bell, and Co.. OCLC 7743994.
- ↑ Seeley, Harry G. (1875). «On the maxillary bone of a new dinosaur (Priodontognathus phillipsii), contained in the Woodwardian Museum of the University of Cambridge». Quarterly Journal of the Geological Society of London 31: pp. 439–443.
- ↑ J. I. Ruiz-Omeñaca. (2011) "Delapparentia turolensis nov. gen. et sp., un nuevo dinosaurio iguanodontoideo (Ornithischia: Ornithopoda) en el Cretácico Inferior de Galve." Estudios Geológicos (advance online publication) doi 10.3989/egeol.40276.124
- ↑ Horner, John R., David B. Weishampel and Catherine A. Forster. "Hadrosauridae". The Dinosauria, pp 438–463.
- ↑ Olshevsky, G. «Dinosaurs of China, Mongolia, and Eastern Asia [under Altirhinus]». Consultado el 22-02-2007.
- ↑ Sauvage, H. E. (1888). «Sur les reptiles trouvés dans le Portlandian supérieur de Boulogne-sur-mer» (en francés). Bulletin du Muséum National d'Historie Naturalle, Paris 3 (16): pp. 626.
- ↑ Upchurch, Paul, Paul M. Barrett, and Peter Dodson. "Sauropoda". The Dinosauria
- ↑ Upchurch, Paul; John Martin (March 2003). «The Anatomy and Taxonomy of Cetiosaurus (Saurischia, Sauropoda) from the Middle Jurassic of England». Journal of Vertebrate Palaeontology 23 (1): pp. 208–231. doi:.
- ↑ Olshevsky, G. «Re: What are these dinosaurs?». Consultado el 16-02-2007.
- ↑ Bakker, R.T. "When Dinosaurs Invented Flowers". The Dinosaur Heresies, 179–198
- ↑ Barrett, Paul M.; Willis, K.J. (2001). «Did dinosaurs invent flowers? Dinosaur-angiosperm coevolution revisited». Biological Reviews 76 (3): pp. 411–447. doi:.
- ↑ Weishampel, D.B.; Barrett, P.M.; Coria, R.A.; Le Loeuff, J.; Xu Xing; Zhao Xijin; Sahni, A.; Gomani, E.M.P. y Noto, C.R. "Dinosaur Distribution". The Dinosauria, 517–606.
- ↑ Goodrich, Samuel Griswold (2004) [1859]. Illustrated Natural History of the Animal Kingdom. University of Michigan Library. ISBN 1418158844.
- ↑ Wright, J.L. (1996). Fossil terrestrial trackways: Preservation, taphonomy, and palaeoecological significance.. University of Bristol. pp. 1–300.
- ↑ Wright, J.L. (1999). «Ichnological evidence for the use of the forelimb in iguanodontians». En David M. Unwin. Cretaceous Fossil Vertebrates. Special Papers in Palaeontology, 60. Palaeontological Association. pp. 209–219. ISBN 0901702676.
- ↑ Coombs Jr., Walter P. (1978). «Theoretical aspects of cursorial adaptations in dinosaurs». Quarterly Review of Biology 53 (4): pp. 393–418. doi:. http://links.jstor.org/sici?sici=0033-5770%28197812%2953%3A4%3C393%3ATAOCAI%3E2.0.CO%3B2-3.
- ↑ Glut, Donald F. (2003). Dinosaurs: The Encyclopedia. 3rd Supplement. Jefferson, North Carolina: McFarland & Company, Inc.. pp. 626. ISBN 0-7864-1166-X.
- ↑ Lapparent, Albert-Félix de (1962). «Footprints of dinosaurs in the Lower Cretaceous of Vestspitsbergen — Svalbard». Arbok Norsk Polarinstitutt, 1960: pp. 13–21.
- ↑ Tagert, E. (1846). «On markings in the Hastings sands near Hastings, supposed to be the footprints of birds». Quarterly Journal of the Geological Society of London 2: pp. 267.
- ↑ Beckles, Samuel H. (1854). «On the ornithoidichnites of the Wealden». Quarterly Journal of the Geological Society of London 10: pp. 456–464.
- ↑ Owen, Richard (1858). «Monograph on the Fossil Reptilia of the Wealden and Purbeck Formations. Part IV. Dinosauria (Hylaeosaurus)». Paleontographical Society Monograph 10: pp. 1–26.
- ↑ «Bird-Footed Iguanodon, 1857». Linda Hall Library of Science, Engineering & TechnologyPaper Dinosaurs 1824–1969. Consultado el 14-02-2007.
- ↑ van Beneden, P.J. (1878). «Sur la découverte de reptiles fossiles gigantesques dans le charbonnage de Bernissart, près de Pruwelz.». Bulletin de l'Institut Royal d'Histoire Naturelle de Belgique 3 (1): pp. 1–19.
- ↑ Smith, Dan (26-02-2001). «A site for saur eyes». New Statesman. http://www.newstatesman.com/200102260031.
- ↑ Lucas, Spencer G. (2000). Dinosaurs: The Textbook. Boston: McGraw-Hill. pp. 13. ISBN 0-07-303642-0.
- ↑ Snider, Mike (29-08-2006). «Godzilla arouses atomic terror». USA Today (Gannett Corporation). http://www.usatoday.com/life/movies/news/2006-08-28-godzilla-dvd_x.htm. Consultado el 21-02-2007.
- ↑ «JPL Small-Body Database Browser: 9941 Iguanodon (1989 CB3)». NASA. Consultado el 10-02-2007.
- ↑ Williams, Gareth. «Minor Planet Names: Alphabetical List». Smithsonian Astrophysical Observatory. Consultado el 10-02-2007.
Enlaces externos
 Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios. Wikimedia Commons alberga contenido multimedia sobre IguanodonCommons.
Wikimedia Commons alberga contenido multimedia sobre IguanodonCommons. Wikiespecies tiene un artículo sobre Iguanodon. Wikispecies
Wikiespecies tiene un artículo sobre Iguanodon. Wikispecies- Mucha más información acerca del Iguanodon
- Excelente diseño e interesantes características del Iguanodon en Yahooligans! (En inglés)
- Ornithopoder - Iguanodon (En sueco)
Categorías:- Dinosaurios del Cretácico
- Dinosaurios europeos
- Fósiles
- Iguanodontia














Wikimedia foundation. 2010.