- Evolución humana
-
Antepasados de Homo sapiens Anterior al género Homo Primeros Homininos
- Australopithecus anamensis
- Australopithecus bahrelghazali
- Australopithecus afarensis
- Australopithecus africanus
- Australopithecus garhi
- Australopithecus sediba
Kenyanthropus
Género Homo - Homo habilis
- Homo rudolfensis
- Homo ergaster
- Homo georgicus
- Homo erectus
- Homo cepranensis
- Homo antecessor
- Homo heidelbergensis
- Homo neanderthaliensis
- Homo floresiensis
- Homo rhodesiensis
- Homo sapiens
- Homo sapiens idaltu
- Homo sapiens sapiens
La evolución humana (u hominización) explica el proceso de evolución biológica de la especie humana desde sus ancestros hasta el estado actual. El estudio de dicho proceso requiere un análisis interdisciplinar en el que se aúnen conocimientos procedentes de ciencias como la genética, la antropología física, la paleontología, la estratigrafía, la geocronología, la arqueología y la lingüística.
El término humano, en el contexto de su evolución, se refiere a los individuos del género Homo. Sin embargo, los estudios de la evolución humana incluyen otros homininos, como Ardipithecus, Australopithecus, etc. Los científicos han estimado que las líneas evolutivas de los seres humanos y de los chimpancés se separaron hace entre 5 y 7 millones de años. A partir de esta separación la estirpe humana siguió ramificándose originando nuevas especies, todas extintas actualmente a excepción del Homo sapiens.
Contenido
Etapas en la evolución humana
Los pre-australopitecinos
Los primeros posibles homínidos bípedos (homininos) son Sahelanthropus tchadiensis (con una antigüedad de 6 ó 7 millones de años), Orrorin tugenensis (unos 6 millones de años) y Ardipithecus (entre 5,5 y 4,5 millones de años). Los fósiles de estos homínidos son escasos y fragmentarios y no hay acuerdo general sobre si eran totalmente bípedos. No obstante, tras el descubrimiento del esqueleto casi completo apodado Ardi, se han podido resolver algunas dudas al respecto; así, la forma de la parte superior de la pelvis indica que era bípedo y que caminaba con la espalda recta, pero la forma del pie, con el dedo gordo dirigido hacia adentro (como en las manos) en vez de ser paralelo a los demás, indica que debía caminar apoyándose sobre la parte externa de los pies y que no podía recorrer grandes distancias.[1]
Los australopitecinos
 Reconstrucción de Australopithecus afarensis.
Reconstrucción de Australopithecus afarensis.
Los primeros homininos de los que se tiene la seguridad de que fueron completamente bípedos son los miembros del género Australopithecus, de los que se han conservado esqueletos muy completos (como el de la famosa Lucy).
Este tipo de hominino prosperó en las sabanas arboladas del este de África entre 4 y 2,5 millones de años atrás con notable éxito ecológico, como lo demuestra la radiación que experimentó, con al menos cinco especies diferentes esparcidas desde Etiopía y el Chad hasta Sudáfrica.
Su desaparición se ha atribuido a la crisis climática que se inició hace unos 2,8 millones de años y que condujo a una desertificación de la sabana con la consiguiente expansión de los ecosistemas abiertos, esteparios. Como resultado de esta presión evolutiva, algunos Australopithecus se especializaron en la explotación de productos vegetales duros y de escaso valor nutritivo, desarrollando un impresionante aparato masticador, originando al Paranthropus; otros Australopithecus se hicieron paulatinamente más carnívoros, originando a los primeros Homo.
Los primeros Homo
 Reconstrucción de Homo habilis
Reconstrucción de Homo habilisNo se sabe con certeza qué especie originó los primeros miembros del género Homo; se han propuesto a A. africanus, A. afarensis y A. garhi, pero no hay un acuerdo general. También se ha sugerido que Kenyanthropus platyops pudo ser el antepasado de los primeros Homo.[2]
Clásicamente se consideran como perteneciente al género Homo los homínidos capaces de elaborar herramientas de piedra. No obstante, esta visión ha sido puesta en duda en los últimos años; por ejemplo, se ha sugerido que Australopithecus ghari, hace 2,5 millones de años fue capaz de fabricar herramientas.[3] Las primeras herramientas eran muy simples y se encuadran en la industria lítica conocida como Olduvayense o Modo 1. Las más antiguas proceden de la región de Afar (Etiopía) y su antigüedad se estima en unos 2,6 millones de años,[4] pero no existen fósiles de homínidos asociados a ellos (ver El artesano olduvayense).
De esta fase se ha descrito dos especies, Homo rudolfensis y Homo habilis, que habitaron África Oriental entre 2,5 y 1,8 millones de años atrás, que a veces se reúnen en una sola. El volumen craneal de estas especies oscila entre 650 y 800 cm³.
El poblamiento de Eurasia
 Reconstrucción de Homo ergaster.
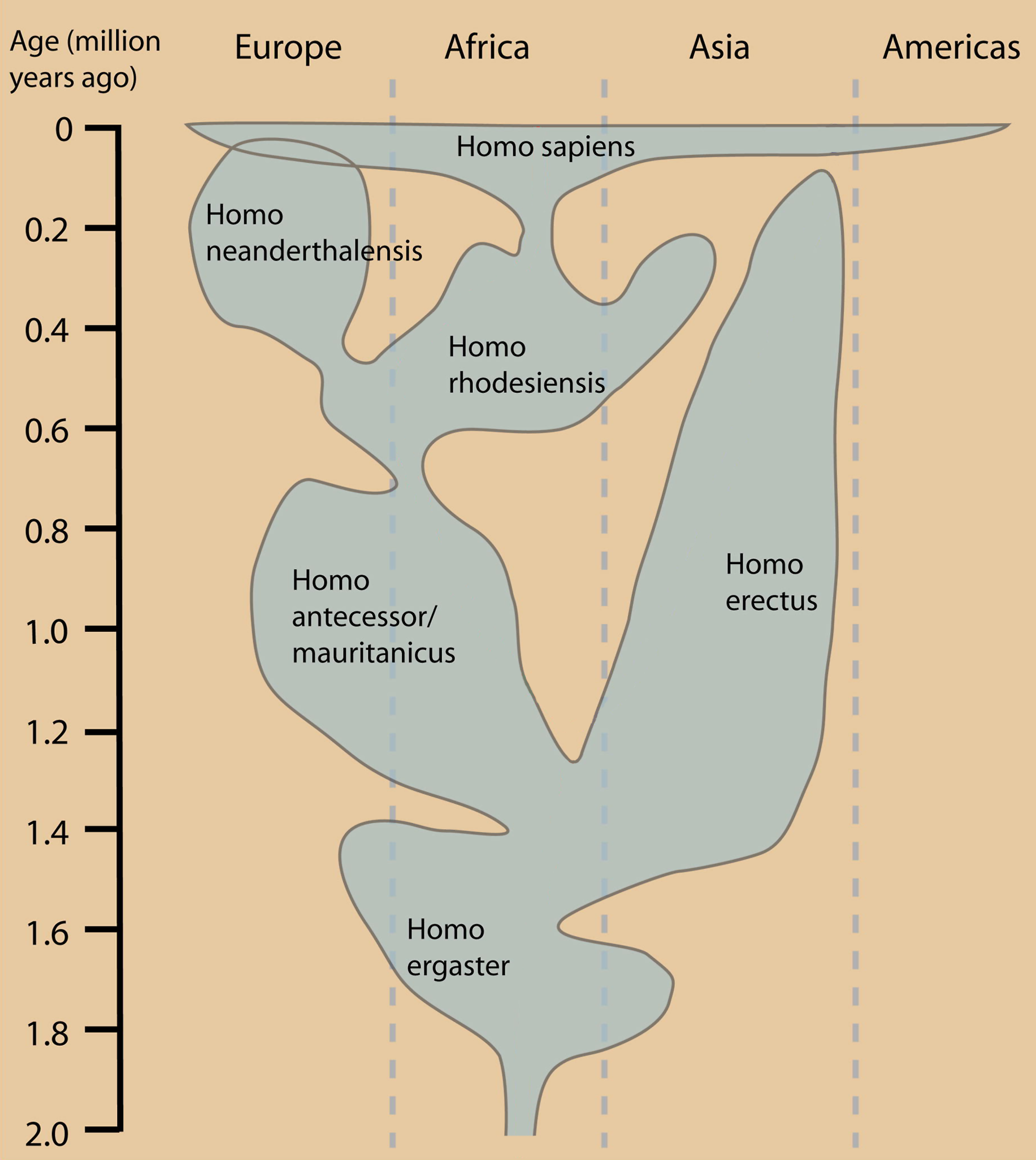
Reconstrucción de Homo ergaster. Distribución geográfica y temporal del género Homo. Otras interpretaciones difieren en la taxonomía y distribución geográfica.
Distribución geográfica y temporal del género Homo. Otras interpretaciones difieren en la taxonomía y distribución geográfica.
Esta es sin duda la etapa más confusa y compleja de la evolución humana. El sucesor cronológico de los citados Homo rudolfensis y Homo habilis es Homo ergaster, cuyos fósiles más antiguos datan de hace aproximadamente 1,8 millones de años, y su volumen craneal oscila entre 850 y 880 cm³.
Morfológicamente es muy similar a Homo erectus y en ocasiones se alude a él como Homo erectus africano. Se supone que fue el primero de nuestros antepasados en abandonar África; se han hallado fósiles asimilables a H. ergaster (o tal vez a Homo habilis) en Dmanisi (Georgia), datados en 1,8 millones de años de antigüedad y que se han denominado Homo georgicus que prueban la temprana salida de África de nuestros antepasados remotos.[5]
Esta primera migración humana condujo a la diferenciación de dos linajes descendientes de Homo ergaster: Homo erectus en Extremo Oriente (China, Java) y Homo antecessor/Homo cepranensis en Europa (España, Italia). Por su parte, los miembros de H. ergaster que permanecieron en África inventaron un modo nuevo de tallar la piedra, más elaborado, denominado Achelense o Modo 2 (hace 1,6 ó 1,7 millones de años). Se ha especulado que los clanes poseedores de la nueva tecnología habrían ocupado los entornos más favorables desplazando a los tecnológicamente menos avanzados, que se vieron obligados a emigrar. Ciertamente sorprende el hecho que H. antecessor y H. erectus siguieran utilizando el primitivo Modo 1 (Olduvayense), cientos de miles de años después del descubrimiento del Achelense. Una explicación alternativa es que la migración se produjera antes de la aparición del Achelense.[6]
Parece que el flujo genético entre las poblaciones africanas, asiáticas y europeas de esta época fue escaso o nulo. Homo erectus pobló Asia Oriental hasta hace solo unos 50.000 años (yacimientos del río Solo en Java) y que pudo diferenciar especies independientes en condiciones de aislamiento, como Homo floresiensis de la Isla de Flores (Indonesia). Por su parte, en Europa se tiene constancia de la presencia humana desde hace casi 1 millón de años (Homo antecessor), pero se han hallado herramientas de piedra más antiguas no asociadas a restos fósiles en diversos lugares. La posición central de H. antecessor como antepasado común de Homo neanderthalensis y Homo sapiens ha sido descartada por los propios descubridores de los restos (Eudald Carbonell y Juan Luis Arsuaga).
Los últimos representantes de esta fase de nuestra evolución son Homo heidelbergensis en Europa, que supuestamente está en la línea evolutiva de los neandertales, y Homo rhodesiensis en África que sería el antepasado del hombre moderno.[7] [8] [9]
Una visión más conservativa de esta etapa de la evolución humana reduce todas las especies mencionadas a una, Homo erectus, que es considerada como una especie politípica de amplia dispersión con numerosas subespecies y poblaciones interfértiles genéticamente interconectadas.
Nuevos orígenes en África
La fase final de la evolución de la especie humana está presidida por tres especies humanas inteligentes, que durante un largo periodo convivieron y compitieron por los mismos recursos. Se trata del Hombre de Neanderthal (Homo neanderthalensis), la especie del homínido de Denisova y el hombre moderno (Homo sapiens). Son en realidad historias paralelas que, en un momento determinado, se cruzan.
El Hombre de Neanderthal surgió y evolucionó en Europa y Oriente Medio hace unos 230.000 años,[6] presentando claras adaptaciones al clima frío de la época (complexión baja y fuerte, nariz ancha).
El homínido de Denisova vivió hace 40.000 años en los montes Altai y probablemente en otras áreas en las cuales también vivieron neandertales y sapiens. El análisis del ADN mitocondrial indica un ancestro femenino común con las otras dos especies hace aproximadamente un millón de años.[10] La secuencia de su genoma ha revelado que habría compartido con los neandertales un ancestro hace unos 650.000 años y con los humanos modernos hace 800.000 años. Un molar descubierto presenta características morfológicas claramente diferentes a las de los neandertales y los humanos modernos.[11]
Los fósiles más antiguos de Homo sapiens datan de hace unos 200.000 años (Etiopía). Hace unos 90.000 años llegó al Próximo Oriente donde se encontró con el Hombre de Neanderthal que huía hacia el sur de la glaciación que se abatía sobre Europa. Homo sapiens siguió su expansión y hace unos 45.000 llegó a Europa Occidental (Francia); paralelamente, el Hombre de Neanderthal se fue retirando, empujado por H. sapiens, a la periferia de su área de distribución (Península ibérica, mesetas altas de Croacia), donde desapareció hace unos 28.000 años.
Aunque H. neanderthalensis ha sido considerado con frecuencia como subespecie de Homo sapiens (H. sapiens neanderthalensis), el análisis del genoma mitocondrial completo de fósiles de H. neanderthalensis sugieren que la diferencia existente es suficiente para considerarlos como dos especies diferentes, separadas desde hace 660.000 (± 140.000) años.[12] (ver el apartado "Clasificación" en Homo neanderthalensis).
Se tiene la casi plena certeza de que el Hombre de Neandertal no es ancestro del ser humano actual, sino una especie de línea evolutiva paralela derivada también del Homo erectus/Homo ergaster a través del eslabón conocido como Homo heidelbergensis. El neandertal coexistió con el Homo sapiens y quizá terminó extinguido por la competencia con nuestra especie. Si existió algún mestizaje entre ambas especies, el aporte a la especie humana actual ha sido, en lo genético, inferior al 5% (un arqueólogo y paleoantropólogo que defendía la hipótesis de una fuerte mixogénesis de las dos especies ha sido descubierto como falsificador de "pruebas"; en efecto, existe actualmente casi total escepticismo de que ambas especies hayan sido interfértiles). En cuanto al llamado Hombre de Cro-Magnon corresponde a las poblaciones de Europa Occidental de la actual especie Homo sapiens.
Homo sapiens
Los parientes vivos más cercanos a nuestra especie son los grandes simios: el gorila, el chimpancé, el bonobo y el orangután.
Demostración palmaria de este parentesco es que un mapeo del genoma humano actual indica que Homo sapiens comparte casi el 99% de los genes con el chimpancé y con el bonobo. Para mayor precisión, el genoma de cualquier individuo de nuestra especie tiene una diferencia de sólo el 0,27% respecto al genoma de Pan troglodytes (chimpancés) y de 0,65% respecto al genoma de los gorilas.
Los fósiles más antiguos de Homo sapiens tienen una antigüedad de casi 200.000 años[13] y proceden del sur de Etiopía (formación Kibish del río Omo), considerada como la cuna de la humanidad (véase Hombres de Kibish). A estos restos fósiles siguen en antigüedad los de Homo sapiens idaltu, con unos 160.000 años.
Biocronología de Hominina 
Algunos datos de genética molecular concordantes con hallazgos paleontológicos, sostienen que todos los seres humanos descienden de una misma Eva mitocondrial o E.M., esto quiere decir que, según los rastreos del ADNmt - que sólo se transmite a través de las madres-, toda la humanidad actual tiene una antecesora común que habría vivido en el noreste de África, probablemente en Tanzania (dada la mayor diversidad genética allí) hace entre 150.000 y 230.000 años[14] (ver haplogrupos de ADN mitocondrial humano).
Estudios de los haplogrupos del cromosoma Y humano, concluyen que por línea paterna hay una ascendencia que llega hasta el Adán cromosómico, el cual habría vivido en el África subsahariana entre hace 60.000 y 90.000 años.[15]
Otros indicios derivados de muy recientes investigaciones sugieren que la de por sí exigua población de Homo sapiens hace unos 74.000 años se redujo al borde de la extinción al producirse el estallido del volcán Toba, según la Teoría de la catástrofe de Toba, volcán ubicado en la isla de Sumatra, cuyo estallido ha dejado como rastro el lago Toba. Tal erupción-estallido tuvo una fuerza 3.000 veces superior a la erupción del Monte Santa Helena en 1980. Esto significó que gran parte del planeta se vio cubierto por nubes de ceniza volcánica que afectaron negativamente a las poblaciones de diversas especies incluidas la humana. Según esta hipótesis llamada entre la comunidad científica Catástrofe de Toba, la población de Homo sapiens (entonces toda en África; la primera migración fuera de África fue en torno al año 70.000 ac) se habría reducido a sólo alrededor de 1000 individuos. Si esto es cierto, significaría que el 'pool' genético de la especie se habría restringido de tal modo que se habría potenciado la unidad genética de la especie humana[cita requerida].
No todos están de acuerdo con esa datación. Después de analizar el ADN de personas de todas las regiones del mundo, el genetista Spencer Wells sostiene que todos los humanos que viven hoy descienden de un solo individuo que vivió en África hace unos 60.000 años.[16]
Por todo lo antedicho queda demostrado el monogenismo de la especie humana y, consecuentemente, descartado el poligenismo, que servía de "argumento" a teorías racistas.
Migraciones prehistóricas de Homo sapiens
Junto a los hallazgos arqueológicos, los principales indicadores de la expansión del ser humano por el planeta son el ADN mitocondrial y el cromosoma Y, que son característicos de la descendencia por línea materna y paterna respectivamente.
Los humanos ya habrían comenzado a salir de África unos 90.000 años antes del presente; colonizando para esas fechas el levante (Estos restos fósiles han sido atribuibles a tempranos Homo sapiens, pero su relación real con los humanos modernos es muy discutible).[17]
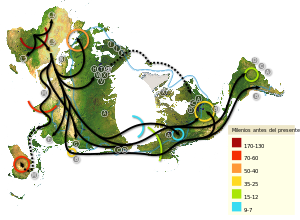 Mapa de la migración humana según estudios del ADN mitocondrial. La leyenda representa los miles de años desde la actualidad. La línea azul señala la extensión máxima de los hielos y las áreas de tundra durante la última gran glaciación.
Mapa de la migración humana según estudios del ADN mitocondrial. La leyenda representa los miles de años desde la actualidad. La línea azul señala la extensión máxima de los hielos y las áreas de tundra durante la última gran glaciación.Australia y Nueva Guinea: la Línea de Wallace no significó para los Homo sapiens un límite insuperable para acceder a esta región. La llegada de humanos a Australia se data hace unos 50.000 años cuando pudieron fabricar rústicas almadías o balsas de juncos para atravesar el estrecho que separaba a Sahul de la región de la Sonda.
Europa: comenzó a ser colonizada hace sólo unos 40.000 años, se supone que durante milenios el desierto de Siria resultaba una barrera infranqueable desde África hacia Europa, por lo que habría resultado más practicable una migración costera desde las costas de Eritrea a las costas yemeníes y de allí al subcontinente indio. La expansión por Europa coincide con la extinción de su coetáneo de entonces, el hombre de Neandertal.
Oceanía: la colonización de estas islas más próximas a Eurasia se habría iniciado hace unos 50.000 años, pero la expansión por esta MUG (macro-unidad geográfica) fue muy lenta y gradual, y hace unos 5.000 años pueblos austronesios comenzaron una efectiva expansión por Oceanía, aunque archipiélagos como el de Hawái y Nueva Zelanda no estaban aún poblados por seres humanos hace 2.000 o 1.500 años (esto requirió el desarrollo de una apropiada técnica naval y conocimientos suficientes de náutica).
América: la llegada del hombre a América, se habría iniciado hace unos 20.000 o, al menos, 15.000 años, aunque no hay consenso al respecto. Durante las glaciaciones el nivel de los océanos desciende al grado que el "Viejo Mundo" y el "Nuevo Mundo" forman un megacontinente unido por el Puente de Beringia.
Antártida: ha sido la última MUG descubierta por el español Gabriel de Castilla (1603), siendo poblada desde 1904, y con población nativa desde 1978 (población chilena).
Bipedestación
Los Homininos, primates bípedos, habrían surgido hace unos 6 ó 7 millones de años en África, cuando dicho continente se encontró afectado por una progresiva desecación que redujo las áreas de bosques y selvas. Como adaptación al bioma de sabana aparecieron primates capaces de caminar fácilmente de modo bípedo y mantenerse erguidos (East Side Story;[18] [19] ) . Más aún, en un medio cálido y con fuerte radiación ultravioleta e infrarroja algunas de las mejores soluciones adaptativas son la marcha bípeda y la progresiva reducción de la capa pilosa, lo que evita el excesivo recalentamiento del cuerpo. Hace 150.000 años el norte de África volvió a sufrir una intensa desertización lo cual significó otra gran presión evolutiva como para que se fijaran los rasgos principales de la especie Homo sapiens.
Para lograr la postura y marcha erecta han tenido que aparecer importantes modificaciones:
- Cráneo. Para permitir la bipedestación, el foramen magnum (u orificio occipital por el cual la médula espinal pasa del cráneo a la raquis) se ha desplazado; mientras en los simios el foramen magnum se ubica en la parte posterior del cráneo, en el Homo sapiens (y en sus ancestros directos) el foramen magnun se ha "desplazado" casi hacia la base del mismo.
- Columna vertebral. La columna vertebral bastante rectilínea en los simios, en el Homo sapiens y en sus ancestros bípedos ha adquirido curvaturas que permiten soportar mejor el peso de la parte superior del cuerpo, tales curvaturas tienen un efecto "resorte". Por lo demás la columna vertebral ha podido erguirse casi 90º a la altura de la pelvis; si se compara con un chimpancé se nota que al carecer este primate de la curva lumbar, su cuerpo resulta empujado hacía adelante por el propio peso. En la raquis humana el centro de gravedad se ha desplazado, de modo que el centro de gravedad de todo el cuerpo se sitúa encima del soporte que constituyen los pies; al tener el Homo sapiens una cabeza relativamente grande el centro de gravedad corporal es bastante inestable (y hace que al intentar nadar, el humano tienda a hundirse "de cabeza")[cita requerida]. Otro detalle; las vértebras humanas son más circulares que las de los simios, esto les permite soportar mejor el peso vertical.
- Pelvis. La pelvis se ha debido ensanchar, lo cual ha sido fundamental en la evolución de nuestra especie. Los huesos ilíacos de la región pelviana en los Homo sapiens (e inmediatos antecesores) "giran" hacia el interior de la pelvis, esto le permite soportar mejor el peso de los órganos al estar en posición erecta. La citada modificación de la pelvis implica una disminución importante en la velocidad posible de la carrera por parte de los humanos. La bipedestación implica una posición de la pelvis, que hace que las crías nazcan "prematuras": en efecto, el parto humano es denominado ventral acodado ya que existe casi un ángulo recto entre la cavidad abdominal y la vagina que en el pubis de la mujer es casi frontal, si en todos los otros mamíferos el llamado canal de parto es muy breve, en cambio en las hembras de Homo sapiens es muy prolongado y sinuoso, esto hace dificultosos los alumbramientos. Como se verá más adelante, esto ha sido fundamental en la evolución de nuestra especie.
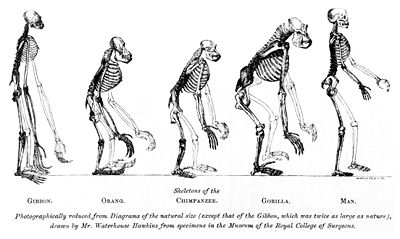
- Piernas. También para la bipedestación ha habido otros cambios morfológicos muy importantes y evidentes, particularmente en los miembros y articulaciones. Los miembros inferiores se han robustecido, el fémur humano se inclina hacia adentro, de modo que le posibilita la marcha sin necesidad de girar casi todo el cuerpo; la articulación de la rodilla se ha vuelto casi omnidireccional (esto es, puede moverse en diversas direcciones), aunque en los monos -por ejemplo el chimpancé- existe una mayor flexibilidad de la articulación de la rodilla, lo que facilita un mejor desplazamiento por las copas de los árboles, es así que el humano a diferencia de sus parientes más próximos no marcha con las rodillas dobladas.
- Pies. En los humanos los pies se han alargado, particularmente en el talón, reduciéndose algo los dedos del pie y dejando de ser oponible el "pulgar" del pie (el dedo mayor), en líneas generales el pie ha perdido casi totalmente la capacidad de aprehensión. Se sabe, en efecto, que el pie humano ha dejado de estar capacitado para aferrarse (cual si fuera una mano) a las ramas, pasando en cambio a tener una función importante en el soporte de todo el cuerpo. El dedo mayor del pie tiene una función vital para lograr el equilibrio de los homininos durante la marcha y la postura erecta; en efecto, el pulgar del pie de un chimpancé es transversal, lo que permite al simio aferrarse más fácilmente de las ramas, en cambio el "pulgar" del pie humano, al estar alineado, facilita el equilibrio y el impulso hacia adelante al marchar o correr. Los huesos de los miembros inferiores son relativamente rectilíneos en comparación con los de otros primates.
Ventajas y desventajas de la bipedestación
Es evidente que la gran cantidad de modificaciones anatómicas que condujeron del cuadrupedismo al bipedismo requirió una fuerte presión selectiva. Se ha discutido mucho sobre la eficacia e ineficacia de la marcha bípeda comparada con la cuadrúpeda. También se ha notado que ningún otro animal de los que se adaptaron a la sabana al final de Mioceno desarrolló una marcha bípeda. Hemos de tener en cuenta que partimos de homínidos con un tipo de desplazamiento cuadrúpedo poco eficaz para largos desplazamientos en terreno abierto: el modo en que se desplazan los chimpancés, apoyando la segunda falange de los dedos de las manos no puede compararse a la marcha cuadrúpeda de ningún otro mamífero. Los primeros homínidos de sabana probablemente se vieron obligados a desplazarse distancias considerables en campo abierto para alcanzar grupos de árboles situados a distancia. La marcha bípeda pudo ser muy eficaz en estas condiciones ya que:[6]
- Permite otear el horizonte por encima de la vegetación herbácea en busca de árboles o depredadores.
- Permite transportar cosas (como comida, palos, piedras o crías) con las manos, liberadas de la función locomotora.
- Es más lenta que la marcha cuadrúpeda, pero es menos costosa energéticamente, lo que debería ser interesante para recorrer largas distancias en la sabana, o en un hábitat más pobre en recursos que la selva.
- Expone menos superficie al sol y permite aprovechar la brisa, lo que ayuda a no recalentar el cuerpo y ahorrar agua, cosa útil en un hábitat con escasez del líquido elemento.
Hace años se argumentó que la liberación de las manos por parte de los primeros homínidos bípedos les permitió elaborar armas de piedra para cazar, lo cual habría sido el principal motor de nuestra evolución. Hoy está claro que la liberación de las manos (que se produjo hace más de 4 millones de años) no está ligada a la fabricación de herramientas, que aconteció unos 2 millones de años después, y que los primeros homininos no eran cazadores y que a lo sumo comían carroña esporádicamente.[cita requerida]
Pero la bipedestación trajo una desventaja en la reproducción, ya que el hecho de pasar del cuadrupedismo al bipedismo conllevó un cambio anatómico de las caderas, con gran reducción del canal del parto que hacia más difícil y doloroso el alumbramiento, tal como se demuestra cuando se compara la cadera de un chimpancé promedio con la de un Australopithecus como Lucy, quienes además presentan un tamaño de cerebro similar.[20]
Aspectos morfológicos
Diferencias con otros primates
Cuando los ancestros del Homo sapiens y otros muchos primates vivían en selvas comiendo frutos, bayas y hojas, abundantes en vitamina C, pudieron perder la capacidad genética, que tiene la mayoría de los animales, de sintetizar en su propio organismo tal vitamina. Tales pérdidas durante la evolución han implicado sutiles pero importantes determinaciones: cuando las selvas originales se redujeron o, por crecimiento demográfico, resultaron superpobladas, los primitivos homininos (y luego los humanos) se vieron forzados a recorrer importantes distancias, migrar, para obtener nuevas fuentes de nutrientes (por ejemplo de la citada vitamina C).
Todos los cambios reseñados han sucedido en un periodo relativamente breve (aunque se mida en millones de años), esto explica la susceptibilidad de nuestra especie a afecciones en la columna vertebral y en la circulación sanguínea y linfática.
Liberación de los miembros superiores
La postura bípeda dejó libres los miembros superiores que ya no tienen que cumplir la función de patas (excepto en los niños muy pequeños) ni la de braquiación, es decir, el desplazamiento de rama en rama con los brazos, aun cuando la actual especie humana, de la cintura hacia arriba mantenga una complexión de tipo arborícola.
Esta liberación de los miembros superiores fue, en su inicio, una adaptación óptima al bioma de sabana; al marchar bípedamente y con los brazos libres, los ancestros del hombre podían recoger más fácilmente su comida; raíces, frutos, hojas, insectos, huevos, reptiles pequeños, roedores y carroña; en efecto, muchos indicios hacen suponer como probable que nuestros ancestros fueran en gran medida carroñeros y, dentro del carroñeo, practicaran la modalidad llamada cleptoparasitismo, esto es, robaban las presas recién cazadas por especies netamente carnívoras; para tal práctica, nuestros ancestros debían haber actuado en bandas, organizadamente.
Los miembros superiores, siempre en relación con otras especies, se han acortado. Estos miembros superiores al quedar liberados de funciones locomotoras, se han podido especializar en funciones netamente humanas. El pulgar oponible es una característica heredada de los primates más antiguos, pero si en éstos la función principal ha sido la de aferrarse a las ramas y en segundo lugar aprehender las frutas o insectos que servían de alimento, en la línea evolutiva que desemboca en nuestra especie la motilidad de la mano, y en particular de los dedos de ésta, se ha hecho gradualmente más precisa y delicada lo que ha facilitado la elaboración de artefactos; aún (junio de 2005) no se tiene conocimiento respecto al momento en que la línea evolutiva comenzó a crear artefactos, es seguro que hace ya más de 2 millones de años Homo habilis/Homo rudolfensis realizaba toscos instrumentos que utilizaba asiduamente (en todo caso, los chimpancés, en estado silvestre, confeccionan "herramientas" de piedra, madera y hueso muy rudimentarias). El desarrollo de la capacidad de pronación en la articulación de la muñeca también ha sido importantísimo para la capacidad de elaborar artefactos.
Visión
El humano hereda de los prosimios la visión estereoscópica y pancromática (la capacidad de ver una amplia tonalidad de los colores del espectro visible); los ojos en la parte delantera de la cabeza posibilitan la visión estereoscópica (en tres dimensiones), pero si esa característica surge en los prosimios como una adaptación para moverse mejor durante la noche o en ambientes umbríos como los de las junglas, en Homo sapiens tal función cobra otro valor; facilita la mirada a lontananza, el otear horizontes, en este aspecto la visión es bastante más aguda en los humanos que en los otros primates y en los prosimios. Esto facilitará el hecho por el cual Homo sapiens sea un ser altamente visual (por ejemplo las comunicaciones mediante la mímica), y facilitará asimismo lo imaginario.
Especialización
Pese al conjunto de modificaciones morfológicas antes reseñadas, desde el punto de vista de la anatomía comparada, llama la atención una cuestión: Homo sapiens es un animal relativamente poco especializado. En efecto, gran parte de las especies animales ha logrado algún tipo de especialización anatómica (por ejemplo los artiodáctilos poseen pezuñas que les permiten correr en las llanuras despejadas), pero las especializaciones, si suelen ser una óptima adaptación a un determinado bioma, conllevan el riesgo de la desaparición de la especie especializada y asociada a tal bioma si éste se modifica.
La ausencia de tales especializaciones anatómicas ha facilitado a los humanos una adaptabilidad inusitada entre las demás especies de vertebrados para adecuarse a muy diversas condiciones ambientales.
Más aún, aunque parezca paradójico, Homo sapiens tiene características neoténicas. En efecto, la estructura craneal de un Homo sapiens adulto se aproxima más a la de la cría de un chimpancé que a la de un chimpancé adulto: el rostro es achatado ("ortognato" o de "bajo índice facial") y es casi inexistente el torus supraorbitario (en la humanidad actual apenas se encuentran vestigios de torus en las poblaciones llamadas australoides). De otro modo se puede decir que los arcos superciliares de Homo sapiens son "infantiles", delicados, el rostro aplanado o ligeramente prognato.
Homo sapiens es, por su anatomía, un animal muy vulnerable si se encuentra en condiciones naturales.
Asociado al hecho por el cual morfológicamente el ser humano tenga características que le aproximan a las de un chimpancé "niño" se encuentra el 'ortognatismo' y esto quiere decir, entre otras cuestiones, que los dientes de Homo sapiens son relativamente pequeños y poco especializados, las mandíbulas, por esto, se ha abreviado y hecho más delicadas, falta además el diastema o espacio en donde encajan los colmillos. La debilidad de las mandíbulas humanas las hace casi totalmente inútiles para la defensa a mordiscos ante un predador y, asimismo, son muy deficientes para poder consumir gran parte del alimento en su estado natural, lo que es uno de los muchos déficits corporales que llevan al humano a vivir en una sociedad organizada.
Tabla comparativa de las diferentes especies del género Homo
- Los nombres en negrita indican la existencia de numerosos registros fósiles.
Especies Cronología (cron) Distribución Altura de adulto (m) Masa de adulto (kg) Volumen craneal (cm³) Registro fósil Descubrimiento /
publicación del nombreH. habilis 2.5–1.4 África oriental 1.0–1.5 30–55 600 Varios 1960/1964 H. rudolfensis 1.9 Kenia 1 cráneo 1972/1986 H. georgicus 1.8–1.6 Georgia 600 Escasos 1999/2002 H. ergaster 1.9–1.25 Este y Sur de África 1.9 700–850 Varios 1975 H. erectus 2–0.3 África, Eurasia (Java, China, Vietnam, Caucaso) 1.8 60 900–1100 Varios 1891/1892 H. cepranensis 0.8 Italia 1 copa craneal 1994/2003 H. antecessor 0.8–0.35 España, Inglaterra 1.75 90 1000 Tres sitios 1994/1997 H. heidelbergensis 0.6–0.25 Europa, África 1.8 60 1100–1400 Varios 1907/1908 Homo rhodesiensis 0.3–0.12 Zambia 1300 Muy pocos 1921 Homo neanderthalensis 0.23–0.024 Europa, Asia Occidental 1.6 55–70 (complexión fuerte) 1200–1700 Varios 1829/1864 Homo sapiens 0.25–presente Mundial 1.4–1.9 55–100 1000–1850 Todavía vive —/1758 H. sapiens idaltu 0.16 Etiopía 1450 3 cráneos 1997/2003 H. floresiensis 0.10–0.012 Indonesia 1.0 25 400 7 individuos 2003/2004 Aparición del lenguaje simbólico
Hablar de la aparición del lenguaje humano, lenguaje simbólico, por lógica parecería implicar que hay que hablar previamente de la cerebración, y eso es bastante cierto, pero el lenguaje humano simbólico tiene sus antecedentes en momentos y cambios morfológicos que son previos a cambios importantes en la estructura del sistema nervioso central. Por ejemplo, los chimpancés pueden realizar un esbozo primario de lenguaje simbólico basándose en la mímica (de un modo semejante a un sistema muy simple de comunicación para sordomudos).
Ahora bien, el lenguaje simbólico por excelencia es el basado en los significantes acústicos, y para que una especie tenga la capacidad de articular sonidos discretos, se requieren más innovaciones morfológicas, algunas de ellas muy probablemente anteriores al desarrollo de un cerebro lo suficientemente complejo como para pensar de modo simbólico. En efecto, observemos la orofaringe y la laringe: en los mamíferos, a excepción del humano, la laringe se encuentra en la parte alta de la garganta, de modo que la epiglotis cierra la tráquea de un modo estanco al beber e ingerir comida. En cambio, en Homo sapiens, la laringe se ubica más abajo, lo que permite a las cuerdas vocales la producción de sonidos más claramente diferenciados y variados, pero al no poder ocluir completamente la epiglotis, la respiración y la ingesta deben alternarse para que el sujeto no se ahogue. El acortamiento del prognatismo que se compensa con una elevación de la bóveda palatina facilitan el lenguaje oral. Otro elemento de relevante importancia es la posición y estructura del hioides, su gracilidad y motilidad permitirán un lenguaje oral lo suficientemente articulado.
Estudios realizados en la Sierra de Atapuerca (España) evidencian que Homo antecessor, hace unos 800.000 años, ya tenía la capacidad, al menos en su aparato fonador, para emitir un lenguaje oral lo suficientemente articulado como para ser considerado simbólico, aunque la consuetudinaria fabricación de utensilios (por toscos que fueran) por parte del Homo habilis hace unos 2 millones de años, sugiere que en éstos ya existía un lenguaje oral articulado muy rudimentario pero lo suficientemente eficaz como para transmitir la suficiente información o enseñanza para la confección de los toscos artefactos.
Además de todas las condiciones recién mencionadas, imprescindibles para la aparición de un lenguaje simbólico, se debe hacer mención de la aparición del gen FOXP2 que resulta básico para la posibilidad de tal lenguaje y del pensamiento simbólico, como se verá a continuación.
Cerebración
La cerebración y la corticalización son temas que requieren, por sí solos, artículos propios, dado el alcance y la importancia de dichos procesos. Aquí importa comentar de lo mínimo indispensable para comprender la evolución humana.
La cerebración tanto como la corticalización son fenómenos biológicos muy anteriores a la aparición de los homínidos, sin embargo en éstos, y en especial en Homo sapiens, la cerebración y la corticalización adquieren un grado superlativo (hasta el punto que Theilard de Chardin enunció una curiosa teoría, la de la noósfera y noogénesis, esto es: teoría del pensar inteligente, que se basa en la evolución del cerebro).
El cerebro de Homo sapiens, en relación a la masa corporal, es uno de los más grandes. Más llamativo es el consumo de energía metabólica (por ejemplo, la producida por la "combustión" de la glucosa) que requiere el cerebro: un 20% de toda la energía corporal, y aun cuando la longitud de los intestinos humanos evidencian los problemas que se le presentan.
En Homo sapiens el volumen oscila entre los 1.200 a 1.400 cm3, siendo el promedio global actual de 1.350 cm3; sin embargo no basta un incremento del volumen, sino cómo se dispone; esto es: cómo está dispuesta la "estructura" del sistema nervioso central y del cerebro en particular. Por término medio, los Homo neanderthalensis pudieron haber tenido un cerebro de mayor tamaño que el de nuestra especie, pero la morfología de su cráneo demuestra que la estructura cerebral era muy diferente: con escasa frente, los neandertalenses tenían poco desarrollados los lóbulos frontales y, en especial, muy poco desarrollada la corteza prefrontal. El cráneo de Homo sapiens no sólo tiene una frente prominente sino que es también más alto en el occipucio (cráneo muy abovedado), esto permite el desarrollo de los lóbulos frontales. De todos los mamíferos, Homo sapiens es el único que tiene la faz ubicada bajo los lóbulos frontales.
Sin embargo, aún más importante para la evolución del encéfalo parecen haber sido las mutaciones en el posicionamiento del esfenoides.
Se ha hecho mención en el apartado dedicado a la aparición del lenguaje articulado de la importancia del gen FOXP2; dicho gen es el encargado del desarrollo de las áreas del lenguaje y de las áreas de síntesis (las áreas de síntesis se encuentran en la corteza cerebral de los lóbulos frontales). El aumento del cerebro y su especialización permitió la aparición de la llamada lateralización, o sea, una diferencia muy importante entre el hemisferio izquierdo y el hemisferio derecho del cerebro. El hemisferio izquierdo tiene desarrollado en su corteza áreas específicas que posibilitan el lenguaje simbólico basado en significantes acústicos: el área de Wernicke y el área de Broca.
Es casi seguro que ya hace 200.000 años los sujetos de la especie Homo sapiens tenían un potencial intelectual equivalente al de la actualidad, pero para que se activara tal potencial tardaron milenios: el primer registro de conducta artística conocido se data hace sólo unos 75.000 años, los primeros grafismos y expresiones netamente simbólicas fuera del lenguaje hablado se datan hace sólo entre 40.000 y 35.000 años. Las primeras escrituras (" memoria segunda" como bien les llamara Roland Barthes) datan de hace entre 5.500 ó 5.000 años, en el Valle del Nilo ó en la Mesopotamia asiática.
Se ha dicho, también líneas antes, que Homo sapiens mantiene características de estructura craneal "primitivas" ya que recuerdan a las de un chimpancé infantil;, en efecto, tal morfología es la que permite tener la frente sobre el rostro y los lóbulos frontales desarrollados.
La cabeza de Homo sapiens, para contener tal cerebro, es muy grande; aún en el feto y en el neonato, razón principal por la cual los partos son difíciles, sumada a la disposición de la pelvis.
Una solución parcial a esto es la heterocronía: el neonato humano está muy incompletamente desarrollado en el momento del parto; puede decirse (con algo de metáfora) que la gestación en el ser humano no se restringe a los ya de por sí prolongados nueve meses intrauterinos, sino que se prolonga extrauterinamente hasta, al menos, los cuatro primeros años; en efecto, el infante está completamente desvalido durante años, tan es así que, que entre los 2 a 4 años es cuando tiene lo suficientemente desarrolladas las áreas visuales del cerebro como para tener una percepción visual de su propio ser (Estadio del espejo descubierto por Jacques Lacan en la década de 1930). Ahora bien, si Homo sapiens tarda mucho en poder tener una percepción plena de su imagen corporal es interesante saber que es uno de los pocos animales que se percibe al ver su imagen reflejada (sólo se nota esta capacidad en bonobos, chimpancés, y si acaso en gorilas, orangutanes, delfines y elefantes).
Tal es la prematuración de Homo sapiens, que mientras un chimpancé neonato tiene una capacidad cerebral de un 65% de la de un chimpancé adulto, o la capacidad de Australopithecus afarensis era en el parto de un 50% respecto a la de su edad adulta, en Homo sapiens 'bebé' tal capacidad no supera al 25% de la capacidad que tendrá a los 45 años (a los 45 años aproximadamente es cuando se desarrolla totalmente el cerebro humano).
Pero no basta el desarrollo cronológico. Para que el cerebro humano se "despliegue" -por así decirlo- o desarrolle requiere de estimulación y afecto; de otro modo la organización de algunas de las áreas del cerebro puede quedar atrofiada.
Cuadro sinóptico de la evolución humana
Época[21] Edad Tiempo (absoluto) Australopitecinos (África) Homo en África Homo en Europa Homo en Asia Cultura Holoceno (reciente) Actualidad
11.700H. sapiens H. sapiens H. sapiens Neolítico a actualidad
(Escritura, ...)Pleistoceno Tarantiense[22] 11.700
126.000H. sapiens
(195.000-act.)H. sapiens
(¿40.000-act.)
H. neanderthalensis
(230.000-29.000)H. sapiens
(42.000-act.)
H. floresiensis
(75.000-13.000)
H. erectus soloensis
(130.000-50.000)Paleolítico Superior
Musteriense
(Pensamiento abstracto, arte)Ioniense[22] 126.000
781.000H. sapiens idaltu
(185.000)
H. sapiens
(195.000-act.)
H. rhodesiensis
(600.000-160.000)H. neanderthalensis
(230.000-29.000)
H. heidelbergensis
(500.000-250.000)H. erectus
(1,8 Ma-250.000)Musteriense
Achelense
(Fuego)Calabriense[22] 781.000
1,8 MaParanthropus robustus
(2,0-1,2 Ma)
P. boisei
(2,3-1,2 Ma)
Australopithecus sediba
(1,95-1,78 Ma)H. ergaster
(1,75-1 Ma)
H. habilis
(1,9-1,6 Ma)H. cepranensis
(800.000)
H. antecessor
(1,2 Ma-780.000)H. erectus
(1,8 Ma-250.000)
H. georgicus
(1,8 Ma)Achelense
OlduvayenseGelasiense[22] 1,8 Ma
2,59 MaA. sediba
(1,95-1,78 Ma)
P. robustus
(2,0-1,2 Ma)
P. boisei
(2,3-1,2 Ma)
P. aethiopicus
(2,6-2,2 Ma)
A. garhi
(2,5 Ma)
A. africanus
(3-2,5 Ma)H. habilis
(1,9-1,6 Ma)
H. rudolfensis
(2,4-1,9 Ma)
Olduvayense
(Industria lítica)Plioceno Piacenziense 2,59 Ma
3,6 MaA. africanus
(3-2,5 Ma)Kenyanthropus platyops
(3,5 Ma)
A. bahrelghazali
(3,58 ± 0,27 Ma)
A. afarensis
(4-2,7 Ma)Zancleense 3,6 Ma
5,33 MaA. afarensis
(4-2,7 Ma)
A. anamensis
(4,2-3,9 Ma)Futuro de la evolución humana
Una línea del pensamiento asegura que la especie humana ha dejado de evolucionar. La razón que plantea es que los avances en la ciencia ahora permiten sobrevivir a personas que de otra forma habrían muerto (eliminación de la presión selectiva), y también permiten una movilidad a nivel global, diluyendo cualquier novedad genética en una población tan grande (eliminación de la deriva genética).[23]
Sin embargo, existen otras posturas que consideran que son precisamente los adelantos tecnológicos los que impulsan actualmente la evolución humana. Por una parte, se ha propuesto que el entorno actual favorece la reproducción de las personas inteligentes, independientemente de su fuerza física o su estado de salud.[23] Además, es posible que la ingeniería genética humana permita seleccionar las características genéticas de la descendencia.[23] Por otra parte, también se ha propuesto que en el futuro la tecnología posibilite a las personas vivir como seres digitales dentro de cuerpos artificiales.[23]
Referencias
Notas
- ↑ Artículo publicado en el periódico La Vanguardia el 2 de octubre de 2009
- ↑ Leakey, M. G.; Spoor, F.; Brown, F. H.; Gatogo, P. N.; Kiarie C., Leakey, L. N. y McDougall, I. (2001). «New hominin genus from eastern Africa shows diverse middle Pliocene lineages». Nature, 410: 433-440
- ↑ Asfaw, B., White, T., Lovejoy, O., Latimer, B., Simpson, S. & Suwa, G., 1999. Australopithecus garhi: a new species of early hominid from Ethiopia. Science, 23(5414): 629-635.
- ↑ Semaw, Sileshi (2000). «[www.indiana.edu/~origins/X-PDF/Semaw2000.pdf The World’s Oldest Stone Artefacts from Gona, Ethiopia: Their Implications for Understanding Stone Technology and Patterns of Human Evolution Between 2.6–1.5 Million Years Ago]». Journal of Archaeological Science 27 (12). doi:. www.indiana.edu/~origins/X-PDF/Semaw2000.pdf.
- ↑ Vekua A., Lordkipanidze D., Rightmire G. P., Agusti J., Ferring R., Maisuradze G. et al., 2002. A new skull of early Homo from Dmanisi, Georgia". Science, 297: 85-89.
- ↑ a b c Arsuaga, J. L. & Martínez, I. 1998. La especie elegida. Ediciones Temas de Hoy, Madrid, 342pp. ISBN 978-84-7880-909-7
- ↑ Manzi, G. (2004). «Human Evolution at the Matuyama-Brunhes Boundary». Evolutionary Anthropology: Issues, News and Reviews 13: 11-24.
- ↑ White, Tim D. et. al. (2003). «Pleistocene Homo sapiens from Middle Awash, Ethiopia». Nature 423 (6491): 742-747.
- ↑ Arsuaga, J. L., Martinez, I., Lorenzo, C., Gracia, A., Munoz, A., Alonso, O. & Gallego, J. (1999) "The human cranial remains from Gran Dolina Lower Pleistocene site (Sierra de Atapuerca, Spain)". Journal of Human Evolution 37: 431-457.
- ↑ Krause, Johannes; Fu, Qiaomei; Good, Jeffrey M.; Viola, Bence; Shunkov, Michael V.; Derevianko, Anatoli P. & Pääbo, Svante (2010), «The complete mitochondrial DNA genome of an unknown hominin from southern Siberia», Nature 464: 894-897.
- ↑ Reich, David; Richard E. Green, et.al. (22 December, 2010) "Genetic history of an archaic hominin group from Denisova Cave in Siberia"; Nature 468 (1012): 1053–1060.
- ↑ Richard E. Green et. al. (2008) "A Draft Sequence of the Neandertal Genome"; Science 328 (5979): 710-722.
- ↑ McDougall, I., Brown, F. H. & Fleagle, J. G., 2005. Stratigraphic placement and age of modern humans from Kibish, Ethiopia. Nature, 433: 733-736. Abstract
- ↑ Soares, Pedro et al 2009, Supplemental Data, Correcting for Purifying Selection: An Improved Human Mitochondrial Molecular Clock. AJHG, Volume 84
- ↑ Mitochondrial Eve and Y-chromosomal Adam The Genetic Genealogist
- ↑ Documentary Redraws Humans' Family Tree(en idioma inglés)
- ↑ The battle over the emergence of modern humans in Eurasia, Chris Brown, 2002, New Archaeology
- ↑ Coppens, Y., 1994. East Side Story: the origin of humankind. Scientific American, 270: 62-69.
- ↑ Esta cuestión es objeto de debate y se proponen diversas alternativas para intentar explicar la bipedestación. Por ejemplo: S. K. S. Thorpe, R. L. Holder, R. H. Crompton (2007) «Origin of Human Bipedalism As an Adaptation for Locomotion on Flexible Branches». Science, 316(5829): 1328-31. Paul O'Higgins, Sarah Elton (2007) «Walking On Trees». Science, 316(5829): 1292-4
- ↑ Sloan, Christopher 2005, "La historia del origen del hombre." National Geographic.
- ↑ Las épocas, edades y colores usados son los establecidos en la «International Sratigraphic Chart» (2009). Consultado el 10 de abril de 2010.
- ↑ a b c d En 2006 se incorporó al Cuaternario la edad o piso Gelasiense (desde el Plioceno al Pleistoceno), bajando por tanto la fecha de inicio del Pleistoceno a los 2,59 Ma y pasando a tener cuatro divisiones en lugar de las tres clásicas (Pleistoceno Inferior, Medio y Superior, ahora Calabriense, Ioniense y Tarantiense respectivamente). Ver p. ej.: Clague, John y Comité Ejecutivo del INQUA (2006). «Open Letter by INQUA Executive Committee». Quaternary Perspectives 16 (1): 158–159. http://www.inqua.org/documents/QP%2016-1.pdf.
- ↑ a b c d «Evolución humana se detuvo por avances de la ciencia: genetistas». La Jornada (25 de agosto de 2011). Consultado el 26 de agosto de 2011.
Bibliografía complementaria
- Guillén-Salazar, F. (2005). Existo, luego pienso: los primates y la evolución de la inteligencia humana. Madrid: Ateles Editores.
- López Moratalla, Natalia (2007). La dinámica de la evolución humana. Eunsa. ISBN 978-84-313-2438-4.
- Morgan Allman, John (2003), El cerebro en evolución. Ariel: Barcelona.
- Stix, Gary (2008): «Huellas de un pasado lejano». Investigación y Ciencia, 384(sept.): 12-19 (Migraciones prehistóricas)
Véase también
 Wikiversidad alberga proyectos de aprendizaje sobre Evolución humana.Wikiversidad
Wikiversidad alberga proyectos de aprendizaje sobre Evolución humana.Wikiversidad- Anexo:Fósiles de la evolución humana
- East Side Story
- Evolución biológica
- Haplogrupos de ADN mitocondrial humano
- Hipótesis del simio acuático
- Historia de la paleoantropología
- Homo
- Origen del hombre
- Paleoantropología
- Paleontología
- Prehistoria
- La odisea de la especie
Enlaces externos
 Wikimedia Commons alberga contenido multimedia sobre Evolución humana. Commons
Wikimedia Commons alberga contenido multimedia sobre Evolución humana. Commons- Becoming Human Trabajo que analiza la evolución humana desde múltiples puntos de vista (en inglés).
- Proyecto de Mapeamiento Genético Genographic.
- Teoría de las Ciencias Humanas (documento número 7 en español, números 5 y 6 en inglés)
- Cómo el homo sapiens salió de África y pobló el mundo PowerPoint con textos básicos en español





Wikimedia foundation. 2010.