- Formicidae
-
 Hormigas
HormigasRango temporal: 130 Ma-0 Ma Cretácico - Reciente 
Hormiga del género Camponotus en Kibaha, Tanzania.Clasificación científica Reino: Animalia Filo: Arthropoda Subfilo: Hexapoda Clase: Insecta Subclase: Pterygota Infraclase: Neoptera Superorden: Endopterygota Orden: Hymenoptera Suborden: Apocrita Superfamilia: Vespoidea Familia: Formicidae
Latreille, 1809Subfamilias - Aenictinae
- Aenictogitoninae
- Agroecomyrmecinae
- Amblyoponinae (incl. «Apomyrminae»)
- Aneuretinae
- Brachymyrmex
- Cerapachyinae
- Dolichoderinae
- Ecitoninae (incl. «Dorylinae» y «Aenictinae»)
- Ectatomminae
- Formicinae
- Heteroponerinae
- Leptanillinae
- Leptanilloidinae
- Martialinae
- Myrmeciinae (incl. «Nothomyrmeciinae»)
- Myrmicinae
- Paraponerinae
- Ponerinae
- Proceratiinae
- Pseudomyrmecinae
Cladograma subfamiliasMartialinae
Amblyoponinae
Paraponerinae
Agroecomyrmecinae
Proceratiinae
Aenictinae
Dorylini
Aenictogitoninae
Aneuretinae
Pseudomyrmecinae
Ectatomminae
Heteroponerinae
Filogenia de las subfamilias existentes.[1] [2]
*Cerapachyinae es parafilético.Los Formícidos (Formicidae), conocidos comúnmente como hormigas, son una familia de insectos sociales que, como las avispas y las abejas, pertenecen al orden de los himenópteros. Las hormigas evolucionaron de antepasados similares a una avispa a mediados del Cretáceo, hace entre 110 y 130 millones de años, diversificándose tras la expansión de las plantas con flor. Son uno de los grupos zoológicos de mayor éxito y en la actualidad están clasificadas más de 12 000 especies, con estimaciones que superan las 14 000, y con unas tendencias actuales que predicen un total de más de 21 000. Se identifican fácilmente por sus antenas en ángulo y su estructura en tres secciones con una estrecha cintura. La rama de la entomología que las estudia se denomina mirmecología.
Forman colonias de un tamaño que se extiende desde unas docenas de individuos predadores que viven en pequeñas cavidades naturales, a colonias muy organizadas que pueden ocupar grandes territorios compuestas por millones de individuos. Estas grandes colonias consisten sobre todo en hembras estériles sin alas que forman castas de «obreras», «soldados» y otros grupos especializados. Las colonias de hormigas también cuentan con algunos machos fértiles y una o varias hembras fértiles llamadas «reinas». Estas colonias son descritas a veces como superorganismos, dado que las hormigas parecen actuar como una entidad única, trabajando colectivamente en apoyo de la colonia.[3]
Han colonizado casi todas las zonas terrestres del planeta; los únicos lugares que carecen de hormigas indígenas son la Antártida y algunas islas remotas o inhóspitas. Las hormigas prosperan en la mayor parte de estos ecosistemas y se calcula que pueden formar el 15-25% de la biomasa de los animales terrestres.[4] Se estima que hay entre mil billones (1015) y diez mil billones (1016) de hormigas viviendo sobre la Tierra. Se considera que su éxito en tantos entornos se debe a su organización social y a su capacidad para modificar hábitats, a su aprovechamiento de los recursos y a su capacidad de defensa. Su prolongada coevolución con otras especies las ha llevado a desarrollar relaciones miméticas, comensales, parásitas y mutualistas.[5]
Sus sociedades se caracterizan por la división del trabajo, la comunicación entre individuos y la capacidad de resolver problemas complejos.[6] Estos paralelismos con las sociedades humanas han sido durante mucho tiempo fuente de inspiración y objeto de numerosos estudios.[a]
Muchas culturas humanas las utilizan como alimento, medicina y como objeto de rituales. Algunas especies son muy valoradas en su papel de agentes de control biológico.[7] Sin embargo, su capacidad de explotar recursos provoca que las hormigas entren en conflicto con los humanos, puesto que pueden dañar cultivos e invadir edificios. Algunas especies, como las hormigas de fuego (género Solenopsis), son consideradas especies invasoras, ya que se han establecido en nuevas áreas donde han sido introducidas casualmente.[8]
Contenido
Etimología
La palabra «hormiga» aparece por primera vez en el siglo XIV y deriva de la palabra latina formīca, que tiene el mismo significado.[9] Tiene el mismo origen que las palabras correspondientes en otras lenguas romances, como por ejemplo formiga (portugués, catalán y gallego), fourmi (francés) y formica (italiano). El nombre de la familia, Formicidae, también deriva del latín formīca.[10]
Taxonomía y evolución
 Hormiga fosilizada en ámbar.
Hormiga fosilizada en ámbar.
La familia Formicidae pertenece al orden de los himenópteros, que también incluye sínfitos, avispas y abejas. Las hormigas evolucionaron de un linaje dentro de las avispas vespoideas. Los análisis filogenéticos sugieren que las hormigas aparecieron a mediados del Cretáceo, hace aproximadamente entre 130 y 110 millones de años. Tras la expansión de las plantas con flor hace unos 100 millones de años, se diversificaron y asumieron una posición ecológica dominante hace unos 60 millones de años.[11] [12] [13] En 1966, E. O. Wilson y sus colegas identificaron los restos fósiles de una hormiga (Sphecomyrma freyi) que vivió en el Cretáceo. El espécimen, atrapado en ámbar, datado como de hace más de 80 millones de años, tiene características tanto de hormigas como de avispas.[14] Sphecomyrma era probablemente una especie recolectora en la superficie, pero algunos estudios sugieren, en base a grupos como Leptanillinae y Martialinae, que las hormigas primitivas probablemente serían depredadoras bajo el suelo.[2]
Durante el período cretácico, unas pocas especies de hormigas primitivas tenían una amplia distribución en el supercontinente Laurasia (el Hemisferio Norte). Eran escasas en comparación con otros insectos, representando aproximadamente el 1% de la población de insectos. Las hormigas se volvieron dominantes tras la radiación adaptativa a principios del Paleógeno. Durante el Oligoceno y el Mioceno ya representaban el 20-40% de todos los insectos encontrados en los principales yacimientos fósiles. De las especies que vivieron en el Eoceno, aproximadamente un género de cada diez sobrevive en la actualidad. Los géneros que sobreviven hoy comprenden el 56% de los géneros encontrados en fósiles de ámbar bálticos (Oligoceno temprano) y el 92% de los géneros en fósiles de ámbar dominicanos (aparentemente Mioceno temprano).[11] [16]
Las termitas, aunque también se las conoce como «hormigas blancas», no son realmente hormigas y pertenecen al orden de los isópteros, por lo que están más estrechamente relacionadas con las cucarachas y las mantis que con las hormigas. El hecho de que las hormigas y las termitas sean ambas eusociales fue motivado por un proceso de convergencia evolutiva.[17] Las hormigas aterciopeladas parecen grandes hormigas, pero realmente son avispas hembras sin alas.[18] [19]
Distribución y diversidad
Región Número de
especies [20]Neotrópico 2162 Neártico 580 Europa 180 África 2500 Asia 2080 Melanesia 275 Australia 985 Polinesia 42 Habitan en todos los continentes excepto la Antártida y algunas grandes islas, como Groenlandia, Islandia y partes de Polinesia.[21] Las islas hawaianas también carecen de especies de hormigas nativas.[22] Ocupan una gran variedad de nichos ecológicos y son capaces de explotar una amplia gama de recursos alimenticios actuando como herbívoras directas o indirectas, depredadoras y carroñeras. La mayor parte de especies son omnívoras generalistas pero algunas se alimentan de forma especializada.
Se estima que hay entre mil billones (1015) y diez mil billones (1016) de hormigas viviendo sobre la Tierra.[23] [24] Su dominio ecológico se puede medir por su biomasa: estimaciones realizadas en distintos entornos indican que representan por término medio el 15-20% del total de la biomasa de los animales terrestres, que se eleva a casi el 25% en la zona tropical.[25] De acuerdo con estas estimaciones, la biomasa de todas las hormigas existentes en el mundo sería similar a la biomasa total de todos los seres humanos.[26]
Su rango de tamaño varía entre 0,75 y 52 mm.[27] [28] La extinta Titanomyrma giganteum es la hormiga gigante de mayor tamaño de la que se tiene conocimiento, mayor incluso que las del género Dorylus, las mayores hormigas gigantes existentes en la actualidad, de unos 5 cm de longitud, que viven en África Oriental y Central;[29] [30] el registro fósil indica que los machos medían unos 3 cm, pero las reinas alcanzaban los 6 cm, con una envergadura de unos 15 cm.[31]
Su color también varía; la mayoría son rojas o negras, el verde es menos habitual, y algunas especies tropicales tienen un tono metálico. Actualmente se conocen más de 12 000 especies, con una estimación superior de unas 14 000 especies y con unas tendencias actuales que predicen un total de de más de 21 000, con la mayor diversidad localizada en la zona tropical.[32] [33] [34] Los estudios taxonómicos continúan desarrollando su clasificación y sistemática, y las bases de datos en línea de especies de hormigas, incluidas AntBase e Hymenoptera Name Server, ayudan a seguir la pista de las especies conocidas y de las descritas más recientemente.[35] La relativa facilidad con la que se pueden recoger especímenes y estudiar las hormigas en los distintos ecosistemas, las ha hecho muy útiles como especie indicadora en estudios de biodiversidad.[36] [37]
Morfología
Tienen unas características morfológicas distintas de otros insectos, como las antenas en codo, glándulas metapleurales y una fuerte constricción de su segundo segmento abdominal en un peciolo en forma de nodo. La cabeza, mesosoma (el tórax más el primer segmento abdominal, fusionado a éste) y metasoma o gáster (el abdomen menos los segmentos abdominales del peciolo) son sus tres segmentos corporales claramente diferenciados. El peciolo forma una cintura estrecha entre su mesosoma y el gáster. El peciolo puede estar formado por uno o dos nodos (sólo el segundo, o el segundo y tercer segmento abdominal).[38]
 Hormiga bulldog mostrando sus potentes mandíbulas y los ojos compuestos relativamente grandes que le proporcionan una visión excelente.
Hormiga bulldog mostrando sus potentes mandíbulas y los ojos compuestos relativamente grandes que le proporcionan una visión excelente.Como el resto de los insectos, las hormigas cuentan con exoesqueleto, una cobertura exterior que sirve de carcasa protectora alrededor del cuerpo y de punto de anclaje para los músculos, en contraste con el endoesqueleto de los humanos y otros vertebrados. Los insectos no tienen pulmones; el oxígeno y otros gases como el dióxido de carbono atraviesan el exoesqueleto a través de unas minúsculas válvulas llamadas espiráculos. Los insectos también carecen de vasos sanguíneos cerrados (sistema circulatorio abierto); en cambio, tienen un tubo perforado, largo y delgado (denominado «aorta dorsal»), que se extiende por la parte superior del cuerpo y que hace las funciones de corazón y bombea hemolinfa hacia la cabeza, gobernando así la circulación de los fluidos internos. El sistema nervioso se compone de un cordón nervioso ventral que se extiende a lo largo del cuerpo, con varios ganglios y ramas que llegan a los extremos de los apéndices.[39]
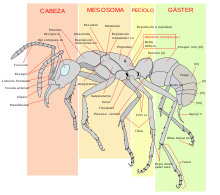 Diagrama de una hormiga obrera.
Diagrama de una hormiga obrera.La cabeza de una hormiga contiene muchos órganos sensoriales. Como la mayor parte de insectos, tienen ojos compuestos formados por numerosas lentes minúsculas unidas. Sus ojos son adecuados para detectar movimiento, pero no ofrecen una gran resolución. También tienen tres pequeños ocelos (ojos simples) en la parte superior de la cabeza, que detectan el nivel lumínico y la polarización de la luz.[40] En comparación con los vertebrados, la mayoría tienen una visión pobre o mediocre, y algunas especies subterráneas son completamente ciegas. Sin embargo, otras especies, como la hormiga bulldog australiana, tienen una vista excepcional. También en la cabeza cuentan con dos antenas, órganos con los que pueden detectar sustancias químicas, corrientes de aire y vibraciones y sirven a su vez para transmitir y recibir señales por medio del tacto. Disponen de dos fuertes mandíbulas, que usan para transportar alimentos, manipular objetos, construir nidos y para defenderse.[39] Algunas especies tienen una cámara intrabucal, una especie de pequeño bolsillo que almacena alimento, para después pasárselo a otras hormigas o a las larvas.[41]
Sus seis patas están ancladas al mesosoma (tórax). Una garra ganchuda situada al final de cada pata ayuda al animal a escalar y a engancharse a varios tipos de superficies. La mayoría de las reinas y los machos tienen alas; las reinas las pierden después del vuelo nupcial, dejando unas marcas visibles que son un rasgo distintivo de las reinas. Sin embargo, en algunas especies las reinas y los machos tampoco tienen alas.[39]
El metasoma (abdomen) de las hormigas alberga órganos internos importantes, incluidos los del sistema reproductor, respiratorio (tráquea) y excretor. Las obreras de muchas especies tienen el ovipositor modificado en un aguijón que usan para someter a las presas y defender sus nidos.[39]
Polimorfismo
 Siete obreras del género Atta de varias castas (izquierda) y dos reinas (derecha).
Siete obreras del género Atta de varias castas (izquierda) y dos reinas (derecha).En las colonias de algunas especies hay castas físicas (con obreras de diferentes clases según el tamaño, denominadas obreras menores, medias y mayores). Es habitual que las hormigas de mayor tamaño tengan cabezas desproporcionadamente grandes y, por lo tanto, unas mandíbulas mayores y más fuertes. Estos ejemplares son denominados a veces hormigas «soldados», porque sus mandíbulas más potentes las hacen más eficaces en el combate, a pesar de que continúan siendo obreras y sus «deberes» no suelen variar significativamente de los de las hormigas menores o medias. En algunas especies no existen las medianas y se aprecia una gran diferencia entre las menores y las mayores.[42] Las hormigas tejedoras (género Oecophylla), por ejemplo, tienen una marcada distribución de tamaño bimodal.[43] [44] Otras especies presentan una variación continua en el tamaño de las obreras. Las obreras más pequeñas de la especie Pheidologeton diversus tienen un peso en seco 500 veces inferior al de sus compañeras de mayor tamaño.[45]
Las obreras no pueden aparearse; sin embargo, debido al sistema haplodiploide de determinación sexual de las hormigas, las obreras de ciertas especies pueden poner huevos no fertilizados que resultan en machos haploides completamente fértiles. El papel de las obreras puede cambiar con la edad y, en algunas especies como las llamadas hormigas melíferas (género Myrmecocystus), un cierto número de obreras jóvenes son alimentadas hasta que su gáster se hincha de forma desproporcionada y sirven como auténticos depósitos vivientes de alimento.[46] Inicialmente se creyó que este polimorfismo en la morfología y el comportamiento de las obreras estaba determinado por factores ambientales, como la nutrición o la acción de las hormonas, que conducían a diferentes tipos de desarrollo; sin embargo, se han detectado diferencias genéticas entre las castas obreras en especies del género Acromyrmex.[47] Estos polimorfismos son causados por cambios genéticos relativamente pequeños; las diferencias en un único gen de Solenopsis invicta pueden determinar si la colonia tendrá una o varias reinas.[48] La especie australiana Myrmecia pilosula tiene un único par de cromosomas (los machos, en su condición de haploides, sólo tienen un cromosoma); esto representa el número de cromosomas más bajo conocido en el mundo animal, lo que las convierte en un interesante tema de estudio en la genética y la biología del desarrollo de los insectos sociales.[49] [50]
Desarrollo y reproducción
 Enjambre de hormigas de la carne (Iridomyrmex purpureus) saliendo del nido.
Enjambre de hormigas de la carne (Iridomyrmex purpureus) saliendo del nido.La vida de una hormiga comienza a partir de un huevo. Si éste es fertilizado, nacerá una hembra (diploide); si no, será un macho (haploide). Este tipo de reproducción, característico de los himenópteros, se llama haplodiploidía.
Los formícidos son insectos holometábolos, que quiere decir que se desarrollan por metamorfosis completa, es decir, el estadio larval pasa por un estadio pupal antes de transformarse en imago. La larva permanece prácticamente inmóvil y es alimentada y cuidada por las obreras. A las larvas se les suministra alimento por trofalaxis, un proceso por el cual una hormiga regurgita la comida líquida almacenada en su buche. Los adultos también comparten de este modo los alimentos almacenados dentro del que podemos denominar «estómago social». Las larvas también pueden recibir alimentos sólidos, como por ejemplo huevos tróficos (no fecundados), trozos de presas, semillas traídas por obreras recolectoras o, en el caso de algunas especies, incluso pueden ser transportadas directamente hasta una presa capturada.
Las larvas pasan una serie de mudas y alcanzan el estadio pupal. La pupa tiene los miembros libres, no unidos al cuerpo como en las crisálidas de mariposa.[51] En algunas especies la diferenciación entre reinas y obreras (ambas son hembras), y entre las diferentes castas de obreras (cuando existen), está influida por la alimentación que reciben las larvas. Las influencias genéticas y el polifenismo por el ambiente de desarrollo son complejos y la determinación de castas sigue siendo objeto de investigación.[52] Las larvas y las pupas tienen que permanecer a temperatura relativamente constante para asegurar un desarrollo adecuado, por lo que son trasladadas a menudo de una cámara de cría a otra dentro de la colonia.[53]
Una nueva obrera pasa los primeros días de su vida adulta cuidando de la reina y de las crías. Posteriormente es promovida a tareas de excavación y de mantenimiento del hormiguero y, más adelante, a defender el hormiguero y recolectar alimento. Estos cambios pueden ser bastante repentinos, y definen lo que se denomina castas temporales. Una posible explicación de esta secuencia son las numerosas bajas que se producen durante la recolección, por lo que resulta un riesgo solo aceptable para las hormigas más viejas, que probablemente morirían pronto de causas naturales.[54] [55]
 Hormigas durante el apareamiento.
Hormigas durante el apareamiento.La mayoría de las especies tienen un sistema en que sólo la reina y las hembras fértiles tienen la capacidad de aparearse. Al contrario de la creencia popular, algunos hormigueros tienen múltiples reinas, mientras que otros pueden existir sin reinas. Las obreras capaces de reproducirse reciben el nombre de «gamergates» y las colonias que carecen de reina son llamadas colonias gamergates.[56] [57] Los machos con alas emergen de las pupas junto con las hembras fértiles, también aladas (aunque algunas especies, como las hormigas guerreras, tienen reinas sin alas), y no hacen nada durante su vida aparte de comer y aparearse. La mayoría de las especies de hormigas son univoltinas, y producen una nueva generación cada año.[58] Durante el periodo de apareamiento, que varía dependiendo de la especie, los machos y hembras alados salen al exterior (generalmente los machos lo hacen antes que las hembras) en el llamado vuelo nupcial. Los machos utilizan señales visuales para buscar un lugar de apareamiento común donde convergen otros machos; entonces secretan unas feromonas para que acudan las hembras. Las hembras de algunas especies se aparean con un solo macho, pero las de otras lo hacen con varios machos.[59] Las hembras que se han apareado buscan después un lugar adecuado para empezar una nueva colonia; allí se arrancan las alas y empiezan a poner los huevos y a cuidarlos. Las hembras almacenan el esperma que obtienen durante su vuelo nupcial para fertilizar de manera selectiva los futuros huevos. Las primeras obreras que nacen son débiles y más pequeñas que las que nacen con posterioridad, pero empiezan a servir a la colonia inmediatamente; amplían el hormiguero, buscan alimentos y cuidan de los otros huevos. En la mayoría de las especies, es así como se forman las colonias. En las especies que tienen varias reinas, una de ellas puede abandonar el hormiguero, junto con algunas obreras, para fundar una nueva colonia en otro lugar.[60]
 Una reina fertilizada comienza a excavar una nueva colonia.
Una reina fertilizada comienza a excavar una nueva colonia.Se han descrito una amplia variedad de estrategias reproductivas en distintas especies de hormigas. Se sabe que las hembras de algunas especies tienen la capacidad de reproducirse asexuadamente por partenogénesis telitoquia,[61] y una especie, Mycocepurus smithii, está compuesta sólo por hembras.[62]
Las colonias de hormigas pueden ser longevas. Las reinas pueden vivir hasta treinta años, mientras que las obreras viven entre uno y tres. Los machos, sin embargo, tienen una vida más efímera, y sólo viven unas pocas semanas.[63] Se estima que las hormigas reina viven hasta cien veces más que los insectos solitarios de un tamaño similar.[64]
Permanecen activas a lo largo de todo el año en la zona tropical, pero, en regiones más frías, sobreviven el invierno en un estado de dormancia o inactividad. Las formas de inactividad son variadas y algunas especies de zonas templadas tienen larvas que entran en estado inactivo (diapausa), mientras que otras, sólo son los adultos los que pasan el invierno en un estado de actividad reducida.[65]
Comportamiento y ecología
Comunicación
 Hormigas tejedoras (género Oecophylla) colaborando para desmembrar a una hormiga roja (las dos situadas a los extremos tiran, mientras que la del medio la corta).
Hormigas tejedoras (género Oecophylla) colaborando para desmembrar a una hormiga roja (las dos situadas a los extremos tiran, mientras que la del medio la corta).Las hormigas se comunican entre ellas por medio de feromonas.[66] Estas señales químicas están más desarrolladas en los formícidos que en otros grupos de himenópteros. Como otros insectos, las hormigas perciben olores con sus largas y delgadas antenas móviles, que ofrecen además información sobre la dirección y la intensidad de los olores. Dado que la mayoría viven en tierra, usan la superficie del suelo para dejar rastros de feromonas que las otras hormigas pueden seguir. En las especies que recolectan en grupos, un recolector que encuentra alimento deja un rastro cuando vuelve al hormiguero; las demás siguen este rastro, y después lo refuerzan cuando vuelven a la colonia con alimentos. Cuando se agota la fuente de alimento ya no van dejando el rastro, y las feromonas se disipan lentamente. Este comportamiento les ayuda a adaptarse a los cambios en su ambiente. Por ejemplo, cuando un camino establecido hacia una fuente de alimento queda bloqueado por un obstáculo, las recolectoras lo abandonan para explorar nuevas rutas. Si una hormiga tiene éxito, deja un nuevo rastro durante su regreso para marcar la ruta más corta. Las mejores rutas son seguidas por más hormigas, reforzando el rastro y encontrando de manera gradual el mejor camino.[67]
Las hormigas no usan las feromonas sólo para dejar rastros. Por ejemplo, una hormiga aplastada libera una feromona de alarma que lleva a las que se encuentren en las proximidades a un frenesí atacante y atrae a más hormigas de otros lugares. Algunas especies incluso usan «feromonas de propaganda» para confundir a las especies enemigas y hacerlas luchar entre sí.[68] Las feromonas son producidas por una gran variedad de estructuras, como la glándula de Dufour, las glándulas venenosas y las del intestino posterior, el pigidio, el recto, el esternón y la tibia posterior.[64] Las feromonas también pueden ser intercambiadas cuando se mezclan con la comida y son traspasadas por trofalaxia, acción que permite transmitir información dentro de la colonia.[69] Esto también les permite determinar a qué grupo de trabajo (por ejemplo, recolección o mantenimiento del hormiguero) pertenecen los otros miembros de la colonia.[70] En las especies con castas de reinas, las obreras empiezan a criar nuevas reinas en la colonia cuando la reina dominante deja de producir una feromona específica.[71]
Algunas hormigas producen sonidos por medio de estridulación (haciendo rozar dos partes del cuerpo), utilizando los segmentos del gáster y las mandíbulas. Los sonidos pueden servir para comunicarse con miembros de la colonia o con otras especies.[72] [73]
Defensa
 Hormiga tejedora en posición de lucha, con las mandíbulas abiertas.
Hormiga tejedora en posición de lucha, con las mandíbulas abiertas.Las hormigas atacan y se defienden mordiendo y, en muchas especies, picando (sólo unas pocas especies poseen aguijón propiamente dicho), a menudo inyectando o rociando productos químicos como el ácido fórmico. Se considera que Paraponera clavata, originaria de América Central y América del Sur, tiene la picadura más dolorosa de cualquier insecto, aunque generalmente no suele resultar fatal para los humanos, y recibe la puntuación más alta en el Schmidt Sting Pain Index.[74] La picadura de la especie Myrmecia pilosula puede llegar a ser letal,[75] pero se ha desarrollado un antisuero.[76] Las hormigas del género Solenopsis son las únicas que tienen un saco de veneno que contiene alcaloides de piperidina.[77] Sus picaduras son dolorosas y pueden ser peligrosas para las personas hipersensibles.[78]
 Para proteger su territorio, las hormigas pueden atacar o defenderse de otras de su misma especie.
Para proteger su territorio, las hormigas pueden atacar o defenderse de otras de su misma especie.Las hormigas del género Odontomachus están equipadas con unas mandíbulas llamadas «mandíbulas-trampa», que se cierran más rápido que cualquier otro apéndice predador del reino animal.[79] Un estudio sobre la especie Odontomachus bauri registró velocidades de entre 126 y 230 km/h, con las mandíbulas cerrándose en 130 microsegundos de media. También se comprobó que estas hormigas usaban sus mandíbulas como una catapulta para expulsar intrusos o para lanzarse ellas mismas hacia atrás para evitar una amenaza.[79] Antes de golpear, la hormiga abre al máximo las mandíbulas y quedan trabadas en esa posición gracias a un mecanismo interno. La energía se almacena en un grueso grupo de músculos y se libera de forma explosiva por la estimulación de unos pelos sensoriales en su interior. Las mandíbulas también permiten movimientos lentos y precisos cuando hay que desarrollar otras tareas como el cuidado de las larvas. Las «mandíbulas-trampa» también se encuentran en los géneros Anochetus, Orectognathus y Strumigenys,[79] así como en algunos miembros de la tribu Dacetini,[80] en lo que es un ejemplo de evolución convergente.
Una especie de hormiga malaya de la superespecie Camponotus cylindricus ha ampliado las glándulas mandibulares que se extienden en su gaster. Cuando se les molesta, las obreras rompen la membrana del gaster, causando un estallido de secreciones que contienen acetofenonas y otros productos químicos que inmovilizan a pequeños insectos atacantes. A causa de esta acción, la obrera muere.[81] La defensa suicida de obreras también se ha registrado en la hormiga brasileña Forelius pusillus donde un pequeño grupo de hormigas abandona la seguridad del nido después de sellar la entrada exterior cada tarde.[82]
 Entrada de hormiguero construida de forma que impida que entre el agua de la lluvia.
Entrada de hormiguero construida de forma que impida que entre el agua de la lluvia.Además de defenderse de los depredadores, tienen que proteger sus colonias de los patógenos. Algunas hormigas obreras se encargan de la higiene de la colonia, y entre sus actividades se incluye eliminar los cadáveres de compañeras muertas (necroforesis).[83] En la especie Atta mexicana se ha identificado el ácido oleico como el compuesto liberado por las hormigas muertas que provoca este comportamiento necroforico,[84] mientras las obreras de Linepithema humile reaccionan a la ausencia de compuestos químicos característicos (dolichodial e iridomyrmecin) presentes en la cutícula de sus compañeras de nido vivas.[85]
La elaborada arquitectura del hormiguero las protege de amenazas naturales como por ejemplo las inundaciones y el sobrecalentamiento.[86] [87] Un caso muy curioso es el de las obreras de Cataulacus muticus, una especie arborícola que vive en los huecos de los troncos, que combaten las inundaciones bebiendo agua dentro del nido y expulsándola al exterior.[88] La pequeña Camponotus anderseni, que construye sus nidos en cavidades de los árboles de los manglares, se ha adaptado de una forma notable a las inundaciones provocadas por las mareas; bloquean la entrada al nido con la cabeza de un soldado y evitan la entrada del agua, y ante la falta de aire puro y el incremento de CO2 en el nido, pueden sobrevivir bajo el agua cambiando a una respiración anaeróbica.[89]
Aprendizaje
Muchos animales pueden aprender comportamientos por imitación, pero es posible que las hormigas sean el único grupo, aparte de los mamíferos, en que se ha observado una enseñanza interactiva. Una recolectora experimentada de Temnothorax albipennis conduce a una compañera inexperta a una fuente de alimento recientemente descubierta por medio del proceso extremadamente lento del llamado «reclutamiento en tándem». La hormiga «alumna» obtiene conocimientos de su «tutora». Tanto la tutora como la alumna reconocen como va el progreso de su compañera, haciendo que la tutora vaya más lenta cuando la alumna se queda atrás, y que acelere cuando la alumna se acerca demasiado.[90]
Experimentos controlados con colonias de Cerapachys biroi sugieren que los individuos pueden elegir su papel en el hormiguero basándose en su experiencia anterior. Una generación entera de obreras idénticas fue dividida en dos grupos en los cuales se controló el éxito en la recolección de alimento. Un grupo era recompensado continuamente con presas, mientras que al otro siempre se lo hacía fracasar. Como resultado, los miembros del grupo con éxito intensificaron su actividad recolectora mientras que el grupo sin éxito salía cada vez menos del nido. Un mes más tarde, los recolectores con éxito continuaban con su papel, mientras que el resto había cambiado para especializarse en el cuidado de las crías.[91]
Construcción de colonias

Muchas especies construyen hormigueros complejos, pero otras son nómadas y no crean estructuras permanentes. Pueden construir colonias subterráneas o construirlas en árboles y otras estructuras naturales o artificiales. Estos nidos pueden encontrarse bajo tierra, bajo piedras o troncos, en el interior de troncos, tallos huecos o incluso bellotas. Los materiales que utilizan para construir el hormiguero generalmente incluyen tierra y materia vegetal.[60] Eligen cuidadosamente el lugar donde construir la colonia; Temnothorax albipennis evita los lugares con hormigas muertas, puesto que esto puede indicar la presencia de parásitos o enfermedades. A la primera señal de amenaza abandonan rápidamente las colonias ya establecidas.[92]
Las hormigas legionarias de América del Sur y las hormigas viajeras de África (género Dorylus) no construyen hormigueros permanentes, sino que van alternando el nomadismo con etapas en que las obreras forman un nido temporal. Las obreras utilizan sus propios cuerpos sujetándose unas a otras, creando así la estructura del nido para proteger a la reina y a las larvas, y lo deshacen posteriormente cuando continúan con su viaje.[93]
Las obreras de las hormigas tejedoras construyen nidos en los árboles uniendo hojas; primero las sujetan por medio de «puentes» de obreras y después hacen que las larvas produzcan seda mientras las mueven por los bordes de las hojas. Se han observado métodos de construcción similares en algunas especies de Polyrhachis.[94]
Alimentación
 Muchas especies de hormigas son carroñeras.
Muchas especies de hormigas son carroñeras. Hormigas melíferas (género Myrmecocystus) almacenando comida en previsión de posibles carencias.
Hormigas melíferas (género Myrmecocystus) almacenando comida en previsión de posibles carencias.La mayor parte de las hormigas son depredadoras generalistas, carroñeras o herbívoras directas o indirectas,[13] pero algunas especies han evolucionado hacia una especialización en los modos de obtener alimentos.
Las hormigas cortadoras de hojas (Atta y Acromyrmex) se alimentan exclusivamente de un hongo que sólo crece dentro de sus colonias. Recogen continuamente hojas que después llevan a la colonia, las cortan en trozos pequeños y las ponen en jardines de hongos. Las obreras se especializan en tareas según su tamaño; las más grandes cortan tallos, las medianas mastican las hojas y las más pequeñas cuidan de los hongos. Estas hormigas son lo bastante sensibles como para reconocer la reacción de los hongos ante diferentes tipos de vegetales, aparentemente detectando señales químicas de los hongos. Si un determinado tipo de hoja es tóxico para el hongo, la colonia ya no recogerá más. Las hormigas se alimentan de unas estructuras producidas por los hongos denominadas gongylidia.[95] Unas bacterias simbióticas que se encuentran en la superficie exterior de las hormigas producen unos antibióticos que eliminan las bacterias que podrían dañar los hongos.[96]
En la especie Leptanilla swani (subfamilia Leptanillinae) la larva alimenta con su propia hemolinfa a la reina mediante glándulas especializadas situadas en su protórax y el tercer segmento abdominal.[97] Este comportamiento es similar al de Adetomyrma venatrix (no relacionada con la anterior), una rara y primitiva especie endémica de Madagascar, conocida como hormiga vampiro u hormiga Drácula, debido a que, en lugar de que las larvas regurgiten alimento como es habitual en la mayor parte de las especies, las obreras y las reinas muerden y perforan la piel de las larvas para alimentarse de los fluidos de su cuerpo. Esta sorprendente forma de alimentarse no causa la muerte de la larva, por lo que se denomina «canibalismo no destructivo».[98]
Orientación
Las hormigas recolectoras recorren distancias de hasta 200 metros desde su nido,[99] y suelen encontrar el camino de regreso gracias a los rastros de olor. Algunas especies salen a buscar el alimento por la noche. Las que salen de día en regiones calurosas y áridas corren el peligro de morir por desecación, de forma que la capacidad de encontrar con mayor rapidez el camino de regreso reduce este riesgo. Así, hormigas diurnas de zonas desérticas del género Cataglyphis como Cataglyphis bicolor, que habita en el desierto del Sahara, se orienta recordando la dirección y la distancia que ha recorrido. Para medir la distancia recorrida utilizan una especie de podómetro interno que lleva la cuenta de los pasos realizados,[100] y también evaluando el movimiento de los objetos en su campo visual,[101] y para la dirección toman como referencia la posición del Sol;[102] integran esta información para encontrar la ruta de retorno más corta posible hasta el nido.[103] Como todas las hormigas, también hacen uso de referencias visuales cuando están disponibles,[104] y utilizan otras señales táctiles y olfativas para orientarse.[105] [106] Algunas especies son capaces de utilizar el campo magnético terrestre para orientarse.[107] Sus ojos compuestos tienen células especializadas que detectan la luz polarizada del Sol, que usan para determinar la dirección;[108] [109] estos detectores de polarización son sensibles a la región ultravioleta del espectro luminoso.[110] En algunas especies de hormigas soldado, un grupo de forrajeras que se separen de la columna principal pueden girarse hasta que la primera hormiga de la fila se une a la última y forman un círculo; de esta forma las obreras siguen girando indefinidamente hasta que mueren por agotamiento.[111]
Locomoción
 Harpegnathos saltator, un tipo de hormiga saltadora.
Harpegnathos saltator, un tipo de hormiga saltadora.Las obreras no tienen alas y las hembras fértiles las pierden tras el vuelo nupcial para fundar su propia colonia. Por lo tanto, a diferencia de sus antepasados véspidos, la mayoría de hormigas se desplazan andando. Algunas especies son capaces de saltar; por ejemplo, Harpegnathos saltator es capaz de efectuar un salto sincronizando la acción de sus pares de patas medio y posterior.[112] Existen también otras especies, como la Cephalotes atratus, llamadas hormigas «planeadoras» (este suele ser un rasgo común en la mayoría de hormigas arborícolas). Las hormigas con esta habilidad son capaces de controlar la dirección de su descenso mientras caen.[113]
Algunas especies pueden formar cadenas para pasar sobre zonas de agua, deslizarse bajo tierra, o a través de espacios entre la vegetación. Otras llegan incluso a crear balsas flotantes que les permiten sobrevivir a las inundaciones. Estas balsas pueden desempeñar un papel importante, dado que permiten a las hormigas colonizar islas.[114] Polyrhachis sokolova, una especie de hormiga que se encuentra en los manglares australianos, puede nadar y vivir en colonias subacuáticas. Como no tienen branquias, estas hormigas respiran gracias a bolsas de aire atrapadas en los hormigueros sumergidos.[115]
Cooperación y competencia
 Iridomyrmex purpureus se alimenta de miel. Las hormigas sociales cooperan y recolectan alimento colectivamente.
Iridomyrmex purpureus se alimenta de miel. Las hormigas sociales cooperan y recolectan alimento colectivamente.No todos los formícidos forman el mismo tipo de sociedades. Las hormigas buldog u hormigas gigantes australianas (género Myrmecia) son unas de las más grandes y basales (primitivas). Como prácticamente todas las hormigas, son eusociales, pero su comportamiento social está poco desarrollado en comparación con otras especies. Cada individuo caza solo, utilizando sus grandes ojos en lugar de sus sentidos químicos para encontrar sus presas.[116] [117]
Algunas especies (como Tetramorium caespitum) atacan y capturan colonias de hormigas vecinas. Otras son menos expansionistas pero igual de agresivas; invaden colonias para robar huevos o larvas, que luego comen o bien crían como obreras esclavas. Entre las que efectúan razias, hay algunas muy especializadas, como por ejemplo las hormigas amazonas (Polyergus), que son incapaces de alimentarse por sí mismas y necesitan obreras capturadas para sobrevivir.[118] [119] Las obreras capturadas de las especies esclavizadas Temnothorax han desarrollado una estrategia contraria, y llegan a destruir hasta las dos terceras partes de las pupas hembras de la especie esclavista Protomognathus americanus, aunque perdonan a los machos (que no participan en las razias de asalto cuando son adultos).[120]
Las hormigas identifican a sus compañeras de colonia por su olor, que proviene de las secreciones de hidrocarburos que cubren su exoesqueleto. Si una hormiga se separa de su colonia original, acaba por perder el olor de su colonia. Cualquier hormiga que entre en un hormiguero sin tener un olor coincidente será atacada.[121]
Algunas especies parásitas entran en las colonias de las hormigas hospedantes y se establecen como parásitos sociales; especies como Strumigenys xenos son totalmente dependientes y no tienen obreras, sino que se alimentan de la comida recogida por sus hospedadoras Strumigenys perplexa.[122] [123] Esta forma de parasitismo se puede observar en muchos géneros de formícidos, pero la hormiga parásita por lo general es una especie estrechamente relacionada con su hospedadora. Las parásitas utilizan una gran variedad de métodos para entrar en el hormiguero del hospedador. Una reina parásita puede entrar en el nido hospedante antes de que eclosionen las primeras larvas, por lo que se establece antes que se desarrolle el olor de la colonia. Otras especies utilizan feromonas para confundir a los hospedantes o para engañarlos, de forma que lleven a la reina parásita dentro del nido. Algunas simplemente se abren paso a la fuerza.[124]
Un conflicto entre sexos de una misma especie se puede observar en algunas especies de hormigas en que los ejemplares fértiles aparentemente combaten para producir descendencia que esté lo más estrechamente relacionada con ellos como sea posible. La forma más extrema implica la producción de descendencia clónica. El extremo del conflicto sexual se observa en Wasmannia auropunctata, donde las reinas producen solamente hijas clónicas diploides por medio de partenogénesis telitoquia, y sólo se producen machos, también clónicos, por medio de un proceso en el que los machos eliminan la contribución materna del huevo diploide, por lo que resultan hijos con un genoma nuclear idéntico al del macho progenitor.[125]
Relación con otros organismos
 La hembra de la araña Myrmarachne plataleoides imita una hormiga tejedora para engañar a posibles predadores.
La hembra de la araña Myrmarachne plataleoides imita una hormiga tejedora para engañar a posibles predadores.Las hormigas tienen relaciones simbióticas con una gran variedad de especies, como otras hormigas, insectos, plantas y hongos. Son la presa de muchos animales e incluso algunos hongos. Algunas especies de artrópodos pasan parte de su vida en hormigueros, bien alimentándose de las hormigas, sus larvas, sus huevos y sus reservas de alimentos, o bien escondiéndose de sus predadores. Estos inquilinos pueden asemejarse mucho en su aspecto a las hormigas. La naturaleza de esta imitación de las hormigas (denominada «mirmecomorfia») varía, y en algunos casos incluye el mimetismo batesiano, en el que el mimetismo reduce el riesgo de depredación. Otros muestran mimetismo wasmaniano, un tipo de mimetismo observado únicamente en inquilinos.[126] [127]
Los pulgones y otros insectos hemípteros secretan un líquido dulce denominado mielada cuando se alimentan de savia. Los azúcares de la mielada son una fuente de alimento con alto contenido energético, que recolectan muchas especies de formícidos.[128] En algunos casos, los pulgones secretan la mielada en respuesta a los golpecitos que les dan con las antenas. Las hormigas, a cambio, mantienen a raya a sus predadores y trasladan a los pulgones de unas zonas de alimentación a otras. Cuando migran a una nueva área, muchas colonias se llevan los pulgones para asegurarse un suministro continuo de mielada. Las hormigas también mantienen cochinillas para recoger su mielada. Estas cochinillas pueden llegar a convertirse en una seria plaga de las piñas si hay hormigas dispuestas a protegerlas de sus enemigos naturales.[129]
Las orugas mirmecófilas de la familia Lycaenidae son reunidas en manadas por las hormigas, que las llevan a alimentarse durante el día y las protegen dentro del hormiguero durante la noche. Las orugas tienen una glándula que secreta mielada cuando les dan masajes. Algunas orugas emiten vibraciones y sonidos que son percibidos por las hormigas.[130] Otras orugas han pasado de mirmecófilas a mirmecófagas: emiten una feromona que hace que las hormigas actúen como si la oruga fuera una de sus propias larvas, entonces las hormigas la llevan al hormiguero, donde la oruga devora sus larvas.[131]
Las hormigas cultivadoras de hongos (tribu Attini), cultivan ciertas especies de hongos de los géneros Leucoagaricus o Leucocoprinus de la familia Agaricaceae. En este mutualismo entre las hormigas y los hongos, cada especie depende de la otra para sobrevivir. La hormiga Allomerus decemarticulatus ha desarrollado una asociación a tres bandas con la planta hospedadora Hirtella physophora (Chrysobalanaceae) y un hongo pegajoso que les sirve para atrapar sus presas de insectos.[132]
 Las incursiones de hormigas guerreras destruyen casi toda la vida animal que se cruza en su camino.
Las incursiones de hormigas guerreras destruyen casi toda la vida animal que se cruza en su camino.Las hormigas guerreras (conocidas popularmente como «marabunta») son nómadas y célebres por sus incursiones o «razias», en las que un enorme número de recolectoras invaden simultáneamente determinadas zonas atacando a sus presas en masa.[133] «Ejércitos» de no menos de 1 500 000 de estas hormigas destruyen casi toda la vida animal que se cruza en su camino.[134] Dorylus spp., conocidas localmente como Siafu,[135] atacan todo lo que encuentran a su paso, incluidos los humanos.[136]
Las hormigas Myrmelachista schumanni crean los llamados «jardines del diablo» matando las plantas circundantes inyectándoles ácido fórmico, dejando únicamente los árboles donde hacen sus nidos (Duroia hirsuta). Esto permite a los árboles multiplicarse y ofrece más lugares para que las hormigas puedan anidar en los troncos de Duroia.[137] Aunque algunas hormigas obtienen néctar de las flores, la polinización por parte de estos insectos es rara.[138] Algunas plantas tienen estructuras especiales de exudación de néctar extrafloral que proporcionan alimento a las hormigas, las cuales a cambio protegen la planta de insectos herbívoros.[139] Especies centroamericanas como por ejemplo el cornezuelo (Acacia cornigera) poseen espinas huecas que albergan colonias de hormigas picadoras (Pseudomyrmex ferruginea) que defienden el árbol de los insectos, mamíferos ramoneadores y enredaderas epifitas. Estudios basados en el marcaje isotópico sugieren que las plantas también obtienen nitrógeno de las hormigas simbióticas.[140] A cambio, las hormigas obtienen alimento de pequeños glóbulos (cuerpos de Belt) ricos en lípidos y proteínas. Otro ejemplo de este tipo de ectosimbiosis es el de los árboles del género Macaranga, que tienen tallos adaptados para alojar colonias de hormigas Crematogaster.
 Hormiga transportando un áfido.
Hormiga transportando un áfido.Muchas especies de árboles tropicales tienen semillas que son dispersadas por las hormigas.[141] La dispersión de semillas por parte de las hormigas (denominada mirmecocoria) está muy extendida y recientes estudios estiman que casi el 9% de todas las especies de plantas tienen este tipo de asociación con las hormigas.[142] [143] Algunas plantas en praderas propensas a los incendios son especialmente dependientes de las hormigas para sobrevivir y extenderse, ya que transportan sus semillas a salvo bajo tierra. Muchas de las semillas dispersadas por las hormigas tienen unas estructuras externas especiales, eleosomas, que son utilizadas como alimento por éstas.[144] En los huevos de los insectos palo se puede observar una convergencia, posiblemente una forma de mimetismo. Estos huevos tienen una estructura comestible similar al eleosoma, de forma que las hormigas los llevan a los hormigueros (lo que ayuda a su dispersión y protección), donde eclosionan y abandonan el nido.[145]
La mayoría de las hormigas son depredadoras y se alimentan y obtienen comida de varios insectos sociales, incluso otras hormigas. Algunas especies se especializan en alimentarse de termitas (Megaponera y Termitopone) mientras que otras especies, como las de la subfamilia Cerapachyinae, se alimentan de otras hormigas.[99] Algunas termitas, como Nasutitermes corniger, forman asociaciones con ciertas especies de formícidos para mantener alejadas a otras especies de hormigas predadoras.[146] La avispa tropical Mischocyttarus drewseni cubre la entrada de su nido con un repelente de hormigas químico.[147] Se cree que muchas avispas tropicales construyen sus nidos en árboles y los cubren para protegerse de las hormigas. Las abejas sin aguijón (Trigona y Melipona) usan defensas químicas contra las hormigas.[99]
Las moscas del Viejo Mundo del género Bengalia (familia de los califóridos) son predadoras de las hormigas y cleptoparásitas, robando las presas o las crías de las mandíbulas de las hormigas adultas.[148] Las hembras sin alas ni patas de la mosca jorobada malaya Vestigipoda myrmolarvoidea viven en los nidos de las hormigas del género Aenictus, y son cuidadas por ellas.[148]
Los hongos de los géneros Cordyceps y Ophiocordyceps infectan a las hormigas, lo que las hace subir por las plantas y que claven sus mandíbulas en el tejido de las mismas. El hongo mata a la hormiga, crece en su cadáver, y produce un cuerpo fructífero. Parece que el hongo altera el comportamiento de las hormigas para facilitar la dispersión de sus esporas en un microhábitat adecuado para el hongo.[149] [150] Los parásitos estrepsípteros también manipulan su comportamiento haciéndolas subir tallos de hierba, ayudando al parásito a encontrar pareja.[151] Un nemátodo (Myrmeconema neotropicum) que infecta a las hormigas de la especie Cephalotes atratus hace que los gásteres negros de las obreras se vuelvan rojos, y modifica el comportamiento de la hormiga haciendo que lo lleven muy alto. Los pájaros confunden estos conspicuos gásteres rojos con frutos maduros, como por ejemplo el del llorón colorado, y se las comen. Los excrementos de los pájaros son recogidos por otras hormigas, que llevan como alimento a las crías, contribuyendo a la expansión del nemátodo.[152]

Las ranas veneno de dardo sudamericanas del género Dendrobates se alimentan principalmente de hormigas, y las toxinas que secretan por la piel pueden tener su origen en estos insectos.[153] Algunos pájaros hormigueros y trepatroncos siguen a hormigas guerreras como Eciton burchellii para alimentarse de los insectos que quedan al descubierto al paso de sus masivas incursiones;[154] [155] por un tiempo este comportamiento fue considerado mutualista, pero estudios más recientes demuestran que en realidad se trata de cleptoparasitismo, puesto que los pájaros roban las presas y las hormigas no solo no se benefician, sino que resultan perjudicadas.[156] Algunas aves tienen un comportamiento peculiar llamado «baño de hormigas» (anting en inglés). Se posan sobre hormigueros, o las cogen para ponérselas en las alas y las plumas; es posible que lo hagan para eliminar ectoparásitos, pero es un comportamiento del que todavía no se sabe demasiado.
Los osos hormigueros, pangolines y varias especies marsupiales australianas tienen adaptaciones especiales para vivir a base de una dieta de hormigas. Estas adaptaciones incluyen lenguas largas y pegajosas para capturarlas y fuertes garras para romper los hormigueros. Se ha comprobado que los osos pardos (Ursus arctos) se alimentan de hormigas, y que aproximadamente el 12%, el 16% y el 4% de su volumen fecal en primavera, verano y otoño, respectivamente, está compuesto por hormigas.[157]
Relación con los humanos
 Las hormigas tejedoras se utilizan como control biológico en plantaciones de cítricos del sur de China.
Las hormigas tejedoras se utilizan como control biológico en plantaciones de cítricos del sur de China.Las hormigas desempeñan múltiples papeles ecológicos que resultan beneficiosos para los humanos, como la eliminación de plagas y la aireación del suelo. Se considera que el uso de hormigas tejedoras en el cultivo de cítricos al sur de China es una de las aplicaciones más antiguas conocidas de control biológico.[7] Por otro lado, las hormigas pueden convertirse en un problema cuando invaden edificios, o causan pérdidas económicas en las actividades agrícolas.[158] [159]
En algunas partes del mundo (principalmente en África y Sudamérica), se utilizan hormigas grandes, especialmente hormigas guerreras, como sutura. Para hacerlo, aprietan los bordes de la herida uno contra otro mientras se aplican las hormigas; éstas muerden con fuerza con sus mandíbulas y en ese momento se les corta el cuerpo, dejando sólo la cabeza y la mandíbula para mantener la herida cerrada.[160] [161] [162]
Algunas especies de la familia Ponerinae poseen un veneno altamente tóxico y potencialmente peligroso, que puede requerir de atención médica. Estas especies incluyen Paraponera clavata (hormiga bala o tocandira) y Dinoponera spp. (falsa tocandira) de América del Sur,[163] así como las Myrmecia de Australia.[164]
En Sudáfrica se utilizan para ayudar en la recolección de rooibos (Aspalathus linearis), arbustos que tienen pequeñas semillas utilizadas para hacer infusiones de hierbas. La planta dispersa mucho sus semillas, lo que hace que sea difícil la recolección manual. Las hormigas recogen estas y otras semillas y las almacenan en el hormiguero, de donde los humanos las pueden recoger todas juntas. Se pueden obtener hasta 200 gramos de semillas de cada hormiguero.[165] [166]
Aunque la mayor parte de especies sobreviven a las tentativas de los humanos de erradicarlas, unas cuantas se encuentran amenazadas. Son sobre todo especies isleñas que han desarrollado características especializadas, como las especies en peligro de extinción Aneuretus simoni de Sri Lanka y Adetomyrma venatrix de Madagascar.[167]
Como alimento
Las hormigas y sus larvas se comen en diferentes partes del mundo. Los huevos de dos especies son la base del plato mexicano conocido como escamoles. Se los considera una forma de caviar de insecto y pueden alcanzar un precio de hasta 90 dólares/kg, al ser estacionales y difíciles de encontrar. En el departamento colombiano de Santander, las hormigas culonas (Atta laevigata) se comen después de ser tostadas vivas.[168]
En zonas de la India y en gran parte de Birmania y Tailandia, se sirve una pasta hecha de una especie de hormiga tejedora (Oecophylla smaragdina) como condimento con el curry.[169] Los huevos y larvas de esta hormiga, así como las hormigas en sí, se utilizan en una ensalada tailandesa, yum (ยำ), en un plato llamado yum khai mod daeng (ยำไข่มดแดง) o ensalada de huevos de hormiga roja, un plato originario de Tailandia del Nordeste. Saville-Kent, en su obra Naturalist in Australia, escribió «La belleza, en el caso de la hormiga tejedora, es más que superficial. Su atractiva transparencia, casi similar a la de las golosinas, fue posiblemente lo que incitó los primeros intentos de consumirla por parte de los humanos». Trituradas en agua, de manera parecida a la limonada, «estas hormigas producen una agradable bebida ácida que es muy apreciada por los nativos del norte de Queensland, e incluso por muchos paladares europeos».[170]
En su obra First Summer in the Sierra, John Muir comenta que los paiute de California se comían los ácidos gásteres de las hormigas carpinteras. Los indios mexicanos se comen las obreras repletas, o almacenes de miel vivientes, de la hormiga melífera (Myrmecocystus mexicanus).[170]
Como plaga
 La minúscula hormiga faraón (Monomorium pharaonis) es una plaga importante en hospitales y edificios de oficinas; puede hacer nidos entre hojas de papel.
La minúscula hormiga faraón (Monomorium pharaonis) es una plaga importante en hospitales y edificios de oficinas; puede hacer nidos entre hojas de papel.Algunas especies de hormigas son consideradas plagas,[171] y, debido a la naturaleza adaptativa de sus colonias, eliminarlas por completo es casi imposible. Por lo tanto, la gestión de plagas se centra en controlar las poblaciones locales, en lugar de intentar eliminar una colonia entera, y la mayor parte de las tentativas para su control son soluciones temporales.
Entre las especies con la consideración de plagas se encuentran Tetramorium caespitum, Anoplolepis gracilipes, Camponotus consobrinus, Monomorium pharaonis, Linepithema humile, Tapinoma sessile, Solenopsis invicta, Myrmica rubra y el género Camponotus. Las poblaciones se controlan por medio de cebos de insecticida, en forma granulada o líquida. Las hormigas recogen el cebo como si fuera comida y lo llevan al hormiguero, donde el insecticida se transmite inadvertidamente a otros miembros de la colonia por trofalaxis. El ácido bórico y el bórax son dos insecticidas habituales, al ser relativamente seguros para los humanos. Se puede esparcir cebo por una zona amplia para controlar especies como Solenopsis invicta, que ocupan grandes áreas. Las colonias de esta especie pueden ser destruidas si se sigue su recorrido hasta el nido y se arroja agua hirviendo en éste para matar a la reina. Esto funciona en aproximadamente el 60% de los casos y requiere unos catorce litros de agua por hormiguero.[172]
Como especies invasoras
Entre los cien peores organismos invasores incluidos en la Base de Datos Global de Especies Invasoras, compilada por la UICN/SSC Invasive Species Specialist Group (ISSG), se encuentran cinco hormigas: Anoplolepis gracilipes, Linepithema humile, Pheidole megacephala, Solenopsis invicta y Wasmannia auropunctata.[173] Las hormigas invasoras tienen gran impacto en los ecosistemas al afectar su composición y sus interacciones ecológicas. Por ejemplo, varían la composición de las hormigas nativas y afectan sus importantes roles como predadoras, carroñeras, herbívoras, detritívoras y granívoras, así como su función como fuente alimenticia de una variedad de especies especializadas en hormigas. Alteran también interacciones especializadas con plantas en la dispersión de semillas, la polinización, la protección de plantas mirmecófilas y con animales como los hemípteros productores de mielatos. Los ecosistemas de islas son especialmente sensibles a las hormigas invasoras, sobre todo en las islas oceánicas donde hay pocas especies de hormigas y las invasoras no encuentran competidores o predadores. Muchos invertebrados nativos pueden allí declinar o incluso extinguirse al no tener adaptaciones defensivas contra las hormigas exóticas.[174]
Como control biológico de plagas
Su utilización por el hombre en el control biológico de plagas es muy antiguo. En Yemen se manejaban las hormigas para disminuir las poblaciones de plagas de las palmas datileras.[175] En China desde la Edad Media los agricultores han regulado plagas en los cítricos con la hormiga tejedora Oecophylla smaragdina, y han controlado algunos lepidópteros barrenadores en las plantaciones de caña de azúcar introduciendo colonias de Tetramorium guineense.[176]
Algunas hormigas exóticas a pesar de ser calificadas como invasoras son usadas en algunas regiones como controladores biológicos de plagas, por ejemplo, Pheidole megacephala en el control del tetuán del boniato en Cuba,[177] Wasmannia auropunctata en el control de plagas de cacaotales en Gabón y Camerún,[178] o para ahuyentar diversos artrópodos herbívoros con Solenopsis invicta o con Linepithema humile en los Estados Unidos.[179] [180]
En la ciencia y la tecnología
Los mirmecólogos las estudian en el laboratorio y en su entorno natural; sus variables y complejas estructuras sociales hacen de estos insectos unos organismos modelo ideales. La visión ultravioleta en las hormigas fue descubierta por Sir John Lubbock en 1881.[181] El estudio de los formícidos ha puesto a prueba hipótesis en ecología y sociobiología y ha sido especialmente importante a la hora de examinar los postulados de teorías de selección de parentesco y estrategias evolutivamente estables.[182] Se pueden estudiar sus colonias criándolas o manteniéndolas temporalmente en hormigueros artificiales, unas estructuras de vidrio especialmente fabricadas para este uso.[183] Se pueden marcar ejemplares con diferentes colores para su estudio y seguimiento.[184]
Las exitosas técnicas utilizadas por las colonias han sido estudiadas en informática y en robótica para producir sistemas distribuidos y tolerantes a fallos para resolver problemas. Este campo de la biomimesis ha encaminado estudios sobre su locomoción, con el resultado de motores de búsqueda basados en su «búsqueda por rastros», sistemas informáticos de almacenamiento tolerante a fallos y algoritmos de red.[185]
Al igual que con otros animales, se han realizado estudios sobre estos insectos para su aplicación en tácticas militares como el swarming. Así, tras una serie de experimentos basados en simulaciones, realizados en un estudio sobre la forma en que las hormigas depositan feromonas para coordinar sus búsquedas de alimento y otras actividades, el Departamento de Defensa de los Estados Unidos descubrió que los algoritmos basados en las feromonas consiguieron impresionantes resultados sobre el control de un grupo de vehículos aéreos no tripulados en un ataque sobre objetivos móviles críticos.[186] Otros estudios sobre estrategia militar animal mostraron que las hormigas se diferencian de las abejas y otros insectos sociales por la utilización del swarming no sólo en la búsqueda de comida o en defensa de la colonia, sino también en guerras de expansión territorial contra otras hormigas. Estas guerras son con frecuencia prolongadas, y de una complejidad operacional que las asemeja a las humanas de modo asombroso.[187]
En la cultura

Las hormigas aparecen a menudo en fábulas e historias para niños, representando el trabajo intenso y el esfuerzo cooperativo. También se las menciona en textos religiosos,[188] [189] como en el libro de los Proverbios de la Biblia, donde son utilizadas como buen ejemplo para los humanos por su trabajo duro y su cooperación. En el Corán, se dice que una hormiga advierte a las demás que vuelvan a casa para evitar ser aplastadas por casualidad por Salomón y su ejército que marcha. Sura 27.18[190] En la fábula atribuida a Esopo, La cigarra y la hormiga, se las utiliza como ejemplo de constancia y trabajo duro, pero que al final tiene recompensa.
En algunas partes de África son consideradas mensajeras de los dioses. A menudo se dice que la picadura de algunas especies de hormigas tiene propiedades curativas. Se afirma que la picadura de algunas especies de Pseudomyrmex alivia la fiebre.[191] En la mitología de algunos pueblos amerindios, como por ejemplo la hopi, son consideradas los primeros animales. Otros grupos utilizan su picadura como prueba de resistencia en ritos de iniciación.[192] [193]
Las sociedades que forman las hormigas siempre han fascinado a los humanos, y se ha escrito sobre ellas tanto humorística como seriamente. Mark Twain escribió sobre estos insectos en su obra A Tramp Abroad (1880).[194] Algunos autores contemporáneos las han utilizado como ejemplo para tratar el tema de la relación entre la sociedad y el individuo, como Robert Frost en su poema Departmental y T. H. White en su novela de fantasía The Once and Future King. La trama de la trilogía de ciencia ficción del entomólogo y escritor francés Bernard Werber, Las hormigas, se divide entre el mundo de las hormigas y el de los humanos; las hormigas y su comportamiento son descritos utilizando el conocimiento científico actual sobre las mismas. En tiempos recientes se han producido dibujos animados como La hormiga atómica y taquilleras películas de animación en 3D con estos insectos como protagonistas como Antz, Bichos, una aventura en miniatura o The Ant Bully. También existe un superhéroe de los años 1960 de Marvel Comics llamado Ant-Man (El hombre hormiga), sobre el que se ha anunciado el estreno de una película en 2012 dirigida por Edgar Wright. El renombrado mirmecólogo E. O. Wilson escribió en 2010 un relato corto, Trailhead, para la revista The New Yorker, que describe la vida y muerte de una hormiga reina y la subida y caída de su colonia, desde un punto de vista de unas hormigas.[195]
Entre finales de la década de 1950 y finales de la década del 1970, las granjas de hormigas eran populares juguetes educativos en los Estados Unidos. Las versiones posteriores usan gel transparente en lugar de tierra, lo que permite mayor visibilidad.[196] A principios de la década de 1990, el videojuego SimAnt, que simulaba una colonia de hormigas, ganó el premio CODiE de 1992 como el «Mejor Programa de Simulación».[197]
El carácter chino para hormiga (chino simplificado: 蚁; tradicional: 蟻; pinyin: yǐ) es una combinación de dos logogramas que puede ser interpretado como «insecto (chino simplificado: 虫; traditional: 蟲; pinyin: chóng) que se comporta correctamente (义; 義; yì)».[198] El carácter de chino tradicional yǐ (蟻) utilizado en la escritura japonesa también comparte esta etimología.[199]
También son la inspiración de muchas criaturas de ciencia ficción, como por ejemplo los insectores (o fórmicos, como se denominan en ediciones más recientes) de El juego de Ender, los insectos de Starship Troopers o las hormigas gigantes de la película Them!. En los juegos de estrategia, las especies inspiradas en las hormigas suelen beneficiarse de un ritmo de producción superior gracias a su mentalidad trabajadora, como es el caso de los Klackons en la serie de juegos Master of Orion o los ChCht de Deadlock II. Todos estos personajes suelen tener una «mente colectiva», creencia errónea habitual sobre las colonias de hormigas.[200]
Véase también
Notas
- ↑ Estos paralelismos entre las sociedades humanas y las colonias de insectos sociales, tales como las hormigas y las abejas, en ningún caso se pueden entender desde el punto de vista de la sociología, pues los rasgos propios constitutivos de la sociedad humana serían impensables en un grupo de hormigas.
Referencias
- ↑ Ward, Philip S (2007). «Phylogeny, classification, and species-level taxonomy of ants (Hymenoptera: Formicidae)» (en inglés, PDF). Zootaxa 1668: pp. 549-563. http://www.mapress.com/zootaxa/2007f/zt01668p563.pdf.
- ↑ a b Rabeling C, Brown JM y Verhaagh M (2008). «Newly discovered sister lineage sheds light on early ant evolution» (en inglés, PDF). PNAS 105: pp. 14913-14917. doi:. PMID 18794530. http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2567467/pdf/zpq14913.pdf.
- ↑ Oster GF, Wilson EO (1978) (en inglés). Caste and ecology in the social insects. Princeton: Princeton University Press. pp. 21-22. ISBN 0691023611. http://books.google.com/books?id=RGE0MwY_NWIC&pg=PA21&dq=isbn:0691023611&hl=es#PPA21,M1.
- ↑ Schultz TR (2000). «In search of ant ancestors» (en inglés). Proceedings of the National Academy of Sciences 97 (26): pp. 14028-14029. doi:. PMID 11106367. http://www.pnas.org/content/97/26/14028.full.
- ↑ Hölldobler y Wilson (1990), p. 471
- ↑ Dicke E, Byde A, Cliff D y Layzell P (2004). A. J. Ispeert, M. Murata y N. Wakamiya. ed (en inglés). Proceedings of Biologically Inspired Approaches to Advanced Information Technology: First International Workshop, BioADIT 2004 LNCS 3141. pp. 364-379.
- ↑ a b Hölldobler y Wilson (1990), pp. 619-629
- ↑ «Pest Notes: Ants (Publication 7411)» (en inglés). University of California, Agriculture and Natural Resources (2007). Consultado el 25 de septiembre de 2011.
- ↑ «Formiga» (en catalán). Enciclopèdia Catalana. http://www.diccionari.cat/lexicx.jsp?GECART=0066217. Consultado el 24 de septiembre de 2011.
- ↑ Simpson DP (1979). Cassell's Latin Dictionary (5ª edición). Londres: Cassell Ltd. ISBN 0-304-52257-0.
- ↑ a b Grimaldi D, Agosti D (2001). «A formicine in New Jersey Cretaceous amber (Hymenoptera: Formicidae) and early evolution of the ants» (en inglés). Proceedings of the National Academy of Sciences 97 (25): pp. 13678-13683. doi:. PMID 11078527. http://www.pnas.org/content/97/25/13678.full.
- ↑ Moreau CS, Bell CD, Vila R, Archibald SB, Pierce NE (2006). «Phylogeny of the ants: Diversification in the Age of Angiosperms» (en inglés). Science 312 (5770): pp. 101-104. doi:. PMID 16601190. http://www.sciencemag.org/content/312/5770/101.abstract.
- ↑ a b Wilson EO, Hölldobler B (2005). «The rise of the ants: A phylogenetic and ecological explanation» (en inglés). Proceedings of the National Academy of Sciences 102 (21): pp. 7411-7414. doi:. PMID 15899976. http://www.pnas.org/content/102/21/7411.full.
- ↑ Wilson E O, Carpenter FM, Brown WL (1967). «The first Mesozoic ants» (en inglés). Science 157: pp. 1038-1040. doi:. PMID 17770424.
- ↑ Brothers DJ (1999). «Phylogeny and evolution of wasps, ants and bees (Hymenoptera, Chrysisoidea, Vespoidea, and Apoidea)» (en inglés). Zoologica Scripta 28: pp. 233-249. doi:.
- ↑ Hölldobler y Wilson (1990), pp. 23–24
- ↑ Thorne, Barbara L (1997). «Evolution of eusociality in termites» (en inglés, PDF). Annual Review of Ecology and Systematics 28: pp. 27–53. doi:. http://www.thornelab.umd.edu/Termite_PDFS/EvolutionEusocialityTermites.pdf.
- ↑ «Order Isoptera - Termites» (en inglés). Iowa State University Entomology (21/1/2006). Consultado el 19 de abril de 2009.
- ↑ «Family Mutillidae - Velvet ants» (en inglés). Iowa State University Entomology (7/6/2007). Consultado el 19 de abril de 2009.
- ↑ Hölldobler y Wilson (1990), p. 4
- ↑ Jones, Alice S. «Fantastic ants - Did you know?» (en inglés). National Geographic Magazine. Consultado el 20 de abril de 2009.
- ↑ Thomas, Philip (2007). «Pest Ants in Hawaii» (en inglés). Hawaiian Ecosystems at Risk project (HEAR). Consultado el 20 de abril de 2009.
- ↑ Brusca RC y Brusca GJ (2005). «XXVI». Invertebrados (2ª edición). Madrid y otros: McGraw-Hill-Interamericana. p. 1005. ISBN 0-87893-097-3.
- ↑ Mark W. Moffett (1/8/2006). «Hormigas, insectos civilizados». National Geographic en español. Archivado desde el original, el 17/8/2010. Consultado el 20 de abril de 2009.
- ↑ Schultz TR (2000). «In search of ant ancestors» (en inglés). Proceedings of the National Academy of Sciences 97 (26): pp. 14028-14029. doi:. PMID 11106367. http://www.pnas.org/content/97/26/14028.full.
- ↑ Holldobler, B y Wilson, EO (2009) (en inglés). The superorganism: the beauty, elegance, and strangeness of insect societies. Nueva York: W. W. Norton. p. 5. ISBN 0393067041.
- ↑ Hölldobler y Wilson (1990), p. 589
- ↑ Shattuck, SO (1999) (en inglés). Australian ants: their biology and identification. Collingwood, Victoria: CSIRO. p. 149. ISBN 0-643-06659-4.
- ↑ Susan Milius. «Giant ants once roamed Wyoming», Science News, 4/5/2011. Consultado el 15 de octubre de 2011 (en inglés).
- ↑ Richard Black. «Giant ants spread in warm climes», BBC News, British Broadcasting Corporation, 3/5/2011. Consultado el 15 de octubre de 2011 (en inglés).
- ↑ Schaal S (2006) (en inglés). Encyclopedia of Life Sciences. doi:. Messel. ISBN 0470016175.
- ↑ Wade, Nicholas. «Taking a Cue From Ants on Evolution of Humans», The New York Times, 15/7/2008. Consultado el 18 de abril de 2009 (en inglés).
- ↑ «Hymenoptera name server. Formicidae species count» (en inglés). Ohio State University. Consultado el 24 de septiembre de 2011.
- ↑ La nueva taxonomía de hormigas. Páginas 45-48 en Fernández, F. «Introducción a las hormigas de la región neotropical». Instituto Humboldt, Bogotá. 2003. http://antbase.org/ants/publications/20973/20973.pdf.
- ↑ Agosti D, Johnson NF (eds.) (2005). «Antbase» (en inglés). American Museum of Natural History. Consultado el 20 de abril de 2009.
- ↑ Agosti D, Majer JD, Alonso JE, Schultz TR (eds.) (2000). Ants: Standard methods for measuring and monitoring biodiversity. Smithsonian Institution Press. http://antbase.org/databases/publications_files/publications_20330.htm. Consultado el 30 de mayo de 2009.
- ↑ Johnson, NF (2007). «Hymenoptera Name Server» (en inglés). Ohio State University. Consultado el 20 de abril de 2009.
- ↑ Borror, Triplehorn y Johnson (1989), p. 737
- ↑ a b c d Borror, Triplehorn y Johnson (1989), pp. 24–71
- ↑ Fent K, Rudiger W (1985). «Ocelli: A celestial compass in the desert ant Cataglyphis» (en inglés). Science 228 (4696): pp. 192-194. doi:. PMID 17779641.
- ↑ Eisner T, Happ GM (1962). «The infrabuccal pocket of a formicine ant: a social filtration device» (en inglés). Psyche 69 (3): pp. 107-116. doi:. http://psyche.entclub.org/69/69-107.html.
- ↑ Wilson, EO (1953). «The origin and evolution of polymorphism in ants» (en inglés). Quarterly Review of Biology 28 (2): pp. 136-156. doi:.
- ↑ Weber, NA (1946). «Dimorphism in the African Oecophylla worker and an anomaly (Hym.: Formicidae)» (en inglés, PDF). Annals of the Entomological Society of America 39: pp. 7-10. http://antbase.org/ants/publications/10434/10434.pdf.
- ↑ «A Fossil Ant Colony: New Evidence of Social Antiquity» (en inglés, PDF). Psyche 71 (2): pp. 93-103. 1964. doi:. http://psyche.entclub.org/pdf/71/71-093.pdf.
- ↑ Moffett MW, Tobin JE (1991). «Physical castes in ant workers: a problem for Daceton armigerum and other ants» (en inglés, PDF). Psyche 98: pp. 283-292. doi:. http://psyche.entclub.org/pdf/98/98-283.pdf.
- ↑ Børgesen LW (2000). «Nutritional function of replete workers in the pharaoh's ant, Monomorium pharaonis (L.)». Insectes Sociaux 47 (2): pp. 141-146. doi:.
- ↑ Hughes WOH, Sumner S, Van Borm S, Boomsma JJ (2003). «Worker caste polymorphism has a genetic basis in Acromyrmex leaf-cutting ants» (en inglés). Proceedings of the National Academy of Sciences 100 (16): pp. 9394-9397. doi:. PMID 12878720.
- ↑ Rossa KG, Kriegera MJB, Shoemaker DD (2003). «Alternative genetic foundations for a key social polymorphism in fire ants» (en inglés). Genetics 165: pp. 1853-1867.
- ↑ Crosland MWJ, Crozier RH (1986). «Myrmecia pilosula, an ant with only one pair of chromosomes» (en inglés). Science 231: p. 1278. doi:. PMID 17839565.
- ↑ Tsutsui ND, Suarez AV, Spagna JC, Johnston JS (2008). «The evolution of genome size in ants» (en inglés). BMC Evolutionary Biology 8 (64): p. 64. doi:. http://www.biomedcentral.com/1471-2148/8/64.
- ↑ Gillott, Cedric (1995) (en inglés). Entomology. Springer. p. 325. ISBN 0306449676.
- ↑ Anderson, K. E.; Linksvayer, T. A. y Smith, C. R. (2008). «The causes and consequences of genetic caste determination in ants (Hymenoptera: Formicidae)» (en inglés). Myrmecological News 11: pp. 119–132. http://myrmecologicalnews.org/cms/images/pdf/volume11/mn11_119-132_non-printable.pdf.
- ↑ Hölldobler y Wilson (1990), pp. 351, 372
- ↑ Traniello JFA (1989). «Foraging strategies of ants» (en inglés). Annual Review of Entomology 34: pp. 191-210. doi:.
- ↑ Sorensen A, Busch TM, Vinson SB (1984). «Behavioral flexibility of temporal sub-castes in the fire ant, Solenopsis invicta, in response to food» (en inglés). Psyche 91: pp. 319-332. doi:. http://psyche.entclub.org/91/91-319.html.
- ↑ Fernández, F. (2003). «Breve introducción a la biología social de las hormigas» (PDF). Introducción a las hormigas de la región Neotropical. Bogotá, Colombia: Instituto de Investigación de Recursos Biológicos Alexander Von Humboldt. p. 92. http://www.bio-nica.info/Biblioteca/Fernandez2003Hormigas05.pdf.
- ↑ Peeters C, Holldobler B (1995). «Reproductive cooperation between queens and their mated workers: The complex life history of an ant with a valuable nest» (en inglés, PDF). Proceedings of the National Academy of Sciences 92: pp. 10977-10979. doi:. PMID 11607589. http://www.pnas.org/content/92/24/10977.full.pdf+html.
- ↑ Taylor, Richard W. (2007). «Bloody funny wasps! Speculations on the evolution of eusociality in ants». En Snelling, R. R., B. L. Fisher, & P. S. Ward (en inglés, PDF). Advances in ant systematics (Hymenoptera: Formicidae): homage to E. O. Wilson – 50 years of contributions. Memoirs of the American Entomological Institute, 80. American Entomological Institute. pp. 580-609. http://antbase.org/ants/publications/21292/21292.pdf.
- ↑ Hölldobler & Wilson (1990)
- ↑ a b Hölldobler y Wilson (1990), pp. 143-179
- ↑ Heinze, Jurgen; Kazuki Tsuji (1995). «Ant reproductive strategies» (en inglés, PDF). Researches on Population Ecology Review 37 (2): pp. 135-149. http://meme.biology.tohoku.ac.jp/POPECOL/RP%20PDF/37(2)/pp.135.pdf.
- ↑ Himler, Anna G.; Eric J. Caldera, Boris C. Baer, Hermógenes Fernández-Marín y Ulrich G. Mueller (2009). «No sex in fungus-farming ants or their crops» (en inglés). Proceedings of the Royal Society B 276 (1667): pp. 2611-2616. doi:. http://rspb.royalsocietypublishing.org/content/early/2009/04/08/rspb.2009.0313.full. Resumen divulgativo – BBC News (16/4/2009).
- ↑ Keller L (1998). «Queen lifespan and colony characteristics in ants and termites» (en inglés). Insectes Sociaux 45 (3): pp. 235-246. doi:.
- ↑ a b Franks NR, Resh VH, Cardé RT (2003) (en inglés). Encyclopedia of Insects. San Diego: Academic Press. pp. 29-32. ISBN 0125869908.
- ↑ Kipyatkov VE (2001). «Seasonal life cycles and the forms of dormancy in ants (Hymenoptera, Formicoidea)» (en inglés, PDF). Acta Societatis Zoologicae Bohemicae 65 (2): pp. 198-217. ISSN 1211-376X. http://www.entomology.bio.pu.ru/personal/kipyatkov/pdf/publ/kipyatkov2001b.pdf.
- ↑ Jackson DE, Ratnieks FL (2006). «Communication in ants» (en inglés). Current Biology 16 (15): pp. R570–R574. doi:. PMID 16890508. http://www.cell.com/current-biology/retrieve/pii/S0960982206018343.
- ↑ Goss S, Aron S, Deneubourg JL, Pasteels JM (1989). «Self-organized shortcuts in the Argentine ant» (en inglés). Naturwissenschaften 76 (12): pp. 579-581. doi:.
- ↑ D'Ettorre P, Heinze J (2001). «Sociobiology of slave-making ants» (en inglés). Acta ethologica 3 (2): pp. 67-82. doi:. http://www.springerlink.com/content/cj1arl0gqb2amw7h/.
- ↑ Detrain C, Deneubourg JL, Pasteels JM (1999) (en inglés). Information processing in social insects. Birkhäuser. pp. 224-227. ISBN 3764357924.
- ↑ Greene MJ, Gordon DM (2007). «Structural complexity of chemical recognition cues affects the perception of group membership in the ants Linephithema humile and Aphaenogaster cockerelli» (en inglés). Journal of Experimental Biology 210 (Pt 5): pp. 897-905. doi:. PMID 17297148. http://jeb.biologists.org/content/210/5/897.full.pdf+html.
- ↑ Hölldobler y Wilson (1990), p. 354
- ↑ Hickling R, Brown RL (2000). «Analysis of acoustic communication by ants» (en inglés). Journal of the Acoustical Society of America 108 (4): pp. 1920-1929. doi:.
- ↑ Roces F, Hölldobler B (1996). «Use of stridulation in foraging leaf-cutting ants: Mechanical support during cutting or short-range recruitment signal?» (en inglés). Behavioral Ecology and Sociobiology 39 (5): p. 293. doi:.
- ↑ Evans, David L. (1990) (en inglés). Insect Defenses: Adaptive Mechanisms and Strategies of Prey and Predators. ISBN 0-88706-896-0. http://books.google.com/books?vid=ISBN0887068960&id=jpxkhS7O0p0C&num=30&pg=PA393&lpg=PA393&dq=%22schmidt,+justin%22&vq=schmidt&sig=F0LL8M4ZolVn5zoy4fwqLxXuvVY. Consultado el 30 de mayo de 2009. «El Schmidt Sting Pain Index (Índice de Dolor de Schmidt) es una escala que mide el dolor relativo causado por picaduras de distintos himenópteros. Es un trabajo del entomólogo Justin O. Schmidt, del Centro Carl Hayden Bee Research.»
- ↑ Clarke PS (1986). «The natural history of sensitivity to jack jumper ants (Hymenoptera: Formicidae: Myrmecia pilosula) in Tasmania» (en inglés). Medical Journal of Australia 145 (11-12): pp. 564-566. PMID 3796365.
- ↑ Brown SGA, Heddle RJ, Wiese MD, Blackman KE (2005). «Efficacy of ant venom immunotherapy and whole body extracts» (en inglés). Journal of Allergy and Clinical Immunology 116 (2): pp. 464-465. doi:.
- ↑ Obin MS, Vander Meer RK (1985). «Gaster flagging by fire ants (Solenopsis spp.): Functional significance of venom dispersal behavior» (en inglés). Journal of Chemical Ecology 11 (12): pp. 1757-1768. doi:.
- ↑ Stafford CT (1996). «Hypersensitivity to fire ant venom» (en inglés). Annals of allergy, asthma, & immunology 77 (2): pp. 87-99. doi:.
- ↑ a b c Patek SN, Baio JE, Fisher BL, Suárez AV (2006). «Multifunctionality and mechanical origins: Ballistic jaw propulsion in trap-jaw ants» (en inglés, PDF). Proceedings of the National Academy of Sciences 103 (34): pp. 12787-12792. doi:. PMID 16924120. http://www.pnas.org/content/103/34/12787.full.pdf.
- ↑ Gronenberg W (1996). «The trap-jaw mechanism in the Dacetine ant Daceton Armigerum and Strumigenys sp» (en inglés, PDF). The Journal of Experimental Biology 199 (9): pp. 2021-2033. http://jeb.biologists.org/content/199/9/2021.full.pdf.
- ↑ Jones, T.H.; Clark, D.A.; Edwards, A.A.; Davidson, D.W.; Spande, T.F. y Snelling, Roy R. (2004). «The Chemistry of Exploding Ants, Camponotus spp. (Cylindricus complex)» (en inglés). Journal of Chemical Ecology 30 (8): pp. 1479-1492. doi:.
- ↑ Tofilski,Adam; Couvillon, MJ;Evison, SEF; Helantera, H; Robinson, EJH; Ratnieks, FLW (2008). «Preemptive Defensive Self-Sacrifice by Ant Workers» (en inglés, PDF). The American Naturalist 172 (5): pp. E239–E243. doi:. http://www.cyf-kr.edu.pl/~rotofils/Tofilski_etal_2008.pdf.
- ↑ Julian GE, Cahan S (1999). «Undertaking specialization in the desert leaf-cutter ant Acromyrmex versicolor» (en inglés). Animal Behaviour 58 (2): pp. 437-442. doi:.
- ↑ López-Riquelme GO, Malo EA, Cruz-López L, Fanjul-Moles ML (2006). «Antennal olfactory sensitivity in response to task-related odours of three castes of the ant Atta mexicana (hymenoptera: formicidae)» (en inglés). Physiological Entomology 31 (4): pp. 353-360. doi:.
- ↑ Choe D-H, Millar JG, Rust MK (2009). «Chemical signals associated with life inhibit necrophoresis in Argentine ants» (en inglés). Proceedings of the National Academy of Sciences 106 (20): pp. 8251-8255. doi:.
- ↑ Tschinkel WR (2004). «The nest architecture of the Florida harvester ant, Pogonomyrmex badius» (en inglés). Journal of Insect Science 4 (21): pp. 1-19. http://insectscience.org/4.21/.
- ↑ Peeters C, Hölldobler B, Moffett M, Musthak Ali TM (1994). «“Wall-papering” and elaborate nest architecture in the ponerine ant Harpegnathos saltator» (en inglés). Insectes Sociaux 41: pp. 211-218. doi:.
- ↑ Maschwitz U, Moog J (2000). «Communal peeing: a new mode of flood control in ants» (en inglés). Naturwissenschaften 87 (12): pp. 563-565. doi:.
- ↑ Nielsen MG, Christian KA (2007). «The mangrove ant, Camponotus anderseni switches to anaerobic respiration in response to elevated CO2 levels» (en inglés). Journal of Insect Physiology 53 (5): pp. 505-508. doi:.
- ↑ Franks NR, Richardson T (2006). «Teaching in tandem-running ants» (en inglés, PDF). Nature 439 (7073): p. 153. doi:. PMID 16407943. http://www.escet.urjc.es/%7Efisica/msanjuan/complejidad/hormigas_nature.pdf. Resumen divulgativo – Scientific American (12/1/2006).
- ↑ Ravary F, Lecoutey E, Kaminski G, Châline N, Jaisson P (2007). «Individual experience alone can generate lasting division of labor in ants» (en inglés). Current Biology 17 (15): pp. 1308-1312. doi:.
- ↑ Franks NR, Hooper J, Webb C, Dornhaus A (2005). «Tomb evaders: house-hunting hygiene in ants» (en inglés). Biology Letters 1 (2): pp. 190-192. doi:.
- ↑ Hölldobler y Wilson (1990), p. 573
- ↑ Robson SK, Kohout RJ (2005). «Evolution of nest-weaving behaviour in arboreal nesting ants of the genus Polyrhachis Fr. Smith (Hymenoptera: Formicidae)» (en inglés). Australian Journal of Entomology 44 (2): pp. 164-169. doi:.
- ↑ Fernández, F. (2003). «Relaciones entre hormigs y plantas:una introducción» (PDF). Introducción a las hormigas de la región Neotropical. Bogotá, Colombia: Instituto de Investigación de Recursos Biológicos Alexander Von Humboldt. p. 174. http://www.bio-nica.info/Biblioteca/Fernandez2003Hormigas10.pdf.
- ↑ Schultz TR (1999). «Ants, plants and antibiotics» (en inglés). Nature 398 (6730): pp. 747-748. doi:. http://www.insecta.ufv.br/Entomologia/ent/disciplina/ban%20160/AULAP/aula2/AntsandBacteria.pdf.
- ↑ The Tree of Life Web Project (21/10/2004). «Leptanillini» (en inglés). Consultado el 30 de mayo de 2009.
- ↑ Fisher, B (2004). Steven M Goodman y Jonathan P Benstead. ed (en inglés, PDF). The Natural History of Madagascar. Chicago y Londres: University of Chicago Press. p. 817. ISBN 0226303063. http://www.antweb.org/madagascar/Fisher2003.pdf.
- ↑ a b c Carrol CR, Janzen DH (1973). «Ecology of foraging by ants» (en inglés). Annual Review of Ecology and Systematics 4: pp. 231-257. doi:.
- ↑ Wittlinger M, Wehner R, Wolf H (2006). «The Ant Odometer: Stepping on Stilts and Stumps» (en inglés). Science 312 (5782): pp. 1965–1967. doi:. http://sun.menloschool.org/~dspence/biology/pdfs/ant_odometer.pdf.
- ↑ Ronacher B y Werner R (1995). «Desert ants Cataglyphis fortis use self-induced optic flow to measure distances travelled» (en inglés). Journal of Comparative Physiology A 177 (1): pp. 21-27. doi:. http://www.zool.uzh.ch/static/research/nb_wehner/literatur/pdf95/wehner19953.pdf.
- ↑ Werner, R. (2003). «Desert ant navigation: how miniature brains solve complex tasks» (en inglés). Journal of Comparative Physiology 189 (8): pp. 579–588. doi:. PMID 12879352. http://www.zool.uzh.ch/static/research/nb_wehner/literatur/pdf03/wehner20038.pdf.
- ↑ Sommer S, Wehner R (2004). «The ant's estimation of distance travelled: experiments with desert ants, Cataglyphis fortis» (en inglés). Journal of Comparative Physiology 190 (1): pp. 1-6. doi:. http://www.springerlink.com/content/bywx5wqjchmh85t2/.
- ↑ Åkesson S, Wehner R (2002). «Visual navigation in desert ants Cataglyphis fortis: are snapshots coupled to a celestial system of reference?» (en inglés, PDF). Journal of Experimental Biology 205: pp. 1971-1978. http://jeb.biologists.org/content/205/14/1971.full.pdf+html.
- ↑ Steck K, Hansson B, Knaden M (2009). «Smells like home: Desert ants, Cataglyphis fortis, use olfactory landmarks to pinpoint the nest» (en inglés). Frontiers in Zoology 6 (5). doi:. http://www.biomedcentral.com/content/pdf/1742-9994-6-5.pdf.
- ↑ Seidl T y Wehner R (2006). «Visual and tactile learning of ground structures in desert ants» (en inglés). Journal of Experimental Biology 209: pp. 3336-3344. doi:. http://jeb.biologists.org/content/209/17/3336.full.
- ↑ Banks AN, Srygley RB (2003). «Orientation by magnetic field in leaf-cutter ants, Atta colombica (Hymenoptera: Formicidae)» (en inglés). Ethology 109 (10): pp. 835-846. doi:. http://si-pddr.si.edu/jspui/bitstream/10088/2096/1/Banks_and_Srygley.pdf.
- ↑ Fukushi T (15 junio 2001). «Homing in wood ants, Formica japonica: use of the skyline panorama» (en inglés). Journal of Experimental Biology 204 (12): pp. 2063-2072. PMID 11441048. http://jeb.biologists.org/content/204/12/2063.full.pdf+html.
- ↑ Wehner R, Menzel R (1969). «Homing in the ant Cataglyphis bicolor» (en inglés). Science 164 (3876): pp. 192-194. doi:. PMID 5774195.
- ↑ Chapman, Reginald Frederick (1998) (en inglés). The Insects: Structure and Function (4ª edición). Cambridge University Press. p. 600. ISBN 0521578906.
- ↑ Delsuc F (2003). «Army Ants Trapped by Their Evolutionary History» (en inglés). Public Library of Science (PLoS) Biology 1 (2): p. e37. doi:.
- ↑ Baroni-Urbani C, Boyan GS, Blarer A, Billen J, Musthak Ali TM (1994). «A novel mechanism for jumping in the Indian ant Harpegnathos saltator (Jerdon) (Formicidae, Ponerinae)» (en inglés). Experientia 50: pp. 63-71. doi:.
- ↑ Yanoviak SP, Dudley R, Kaspari M (2005). «Directed aerial descent in canopy ants» (en inglés, PDF). Nature 433 (7026): pp. 624-626. doi:. http://www.canopyants.com/Nature05.pdf.
- ↑ Morrison LW (1998). «A review of Bahamian ant (Hymenoptera: Formicidae) biogeography» (en inglés). Journal of Biogeography 25 (3): pp. 561-571. doi:.
- ↑ Clay RE, Andersen AN (1996). «Ant fauna of a mangrove community in the Australian seasonal tropics, with particular reference to zonation» (en inglés). Australian Journal of Zoology 44: pp. 521-533. doi:.
- ↑ Crosland MWJ, Crozier RH, Jefferson E (1988). «Aspects of the biology of the primitive ant genus Myrmecia F. (Hymenoptera: Formicidae)» (en inglés). Australian Journal of Entomology 27 (4): pp. 305-309. doi:. http://onlinelibrary.wiley.com/doi/10.1111/j.1440-6055.1988.tb01179.x/pdf.
- ↑ Moffett MW (mayo 2007). «Ant, Bulldog Ants» (en inglés). National Geographic. Consultado el 28 de abril de 2009.
- ↑ Ellen Zimmerli y Howard Topoff (1994). «Queens of the socially parasitic antPolyergus do not kill queens ofFormica that have not formed colonies (Hymenoptera: Formicidae)» (en inglés). Journal of Insect Behavior 7 (1): pp. 119-121. doi:. http://www.springerlink.com/content/v55r6w2661041837/fulltext.pdf?page=1.
- ↑ Diehl E, Junqueira LK, Berti-Filho E (2005). «Ant and termite mound coinhabitants in the wetlands of Santo Antonio da Patrulha, Rio Grande do Sul, Brazil». Brazilian Journal of Biology 65 (3): pp. 431-437. doi:. http://www.scielo.br/scielo.php?script=sci_arttext&pid=S1519-69842005000300008&lng=en&nrm=iso.
- ↑ Achenbach A; Foitzik S (2009). «First evidence for slave rebellion: enslaved ant workers systematically kill the brood of their social parasite Protomognathus americanus» (en inglés). Evolution 63 (4): pp. 1068-1075. PMID 19243573.
- ↑ Henderson G, Andersen JF, Phillips JK, Jeanne RL (2005). «Internest aggression and identification of possible nestmate discrimination pheromones in polygynous ant Formica montana» (en inglés). Journal of Chemical Ecology 16 (7): pp. 2217-2228. doi:.
- ↑ Ward PS (1996). «A new workerless social parasite in the ant genus Pseudomyrmex (Hymenoptera: Formicidae), with a discussion of the origin of social parasitism in ants» (en inglés). Systematic Entomology 21: pp. 253-263. doi:. http://www.archive.org/details/ants_08424.
- ↑ Taylor RW (1968). «The Australian workerless inquiline ant, Strumigenys xenos Brown (Hymenoptera-Formicidae) recorded from New Zealand» (en inglés). New Zealand Entomologist 4 (1): pp. 47-49. http://www.archive.org/details/ants_10687.
- ↑ Hölldobler y Wilson (1990), pp. 436-448
- ↑ Fournier, D, Estoup A, Orivel J, Foucaud J, Jourdan H, Le Breton J, Keller L (2005). «Clonal reproduction by males and females in the little fire ant» (en inglés). Nature 435 (7046): pp. 1230-1234. doi:.
- ↑ Reiskind J (1977). «Ant-mimicry in Panamanian clubionid and salticid spiders (Araneae: Clubionidae, Salticidae)» (en inglés). Biotropica 9 (1): pp. 1-8. doi:.
- ↑ Cushing PE (1997). «Myrmecomorphy and myrmecophily in spiders: A Review» (en inglés, PDF). The Florida Entomologist 80 (2): pp. 165-193. doi:. http://www.fcla.edu/FlaEnt/fe80p165.pdf.
- ↑ Styrsky JD, Eubanks MD (enero 2007). «Ecological consequences of interactions between ants and honeydew-producing insects» (en inglés). Proceedings of the Royal Society B. Biological Sciences 274 (1607): pp. 151–164. doi:. PMID 1685857. http://rspb.royalsocietypublishing.org/content/274/1607/151.full?sid=7df3cd77-4b28-48c0-847a-ec3b699ad4b4.
- ↑ Jahn GC, Beardsley JW (1996). «Effects of Pheidole megacephala (Hymenoptera: Formicidae) on survival and dispersal of Dysmicoccus neobrevipes (Homoptera: Pseudococcidae)» (en inglés). Journal of Economic Entomology 89: pp. 1124-1129.
- ↑ DeVries PJ (1992). «Singing caterpillars, ants and symbiosis» (en inglés). Scientific American 267 (4): pp. 76-82. doi:.
- ↑ Pierce NE, Braby MF, Heath A, et ál (2002). «The ecology and evolution of ant association in the Lycaenidae (Lepidoptera)» (en inglés). Annual Review of Entomology 47: pp. 733-771. doi:.
- ↑ Dejean A, Solano PJ, Ayroles J, Corbara B, Orivel J (2005). «Arboreal ants build traps to capture prey» (en inglés). Nature 434 (7036): p. 973. doi:.
- ↑ Alex Chadwick (25/7/2006). «The Savage, Beautiful World of Army Ants» (en inglés). National Public Radio (NPR). Consultado el 1 de noviembre de 2009.
- ↑ «Hymenopteran» (en inglés). Encyclopædia Britannica. Consultado el 1 de noviembre de 2009.
- ↑ «Driver ant, locally known as 'Siafu'» (en inglés). BBC - Science & Nature. Consultado el 1 de noviembre de 2009.
- ↑ «Driver ant» (en inglés). Encyclopædia Britannica. Consultado el 1 de noviembre de 2009.
- ↑ Frederickson ME, Gordon DM (2007). «The devil to pay: a cost of mutualism with Myrmelachista schumanni ants in ‘devil’s gardens’ is increased herbivory on Duroia hirsuta trees» (en inglés, PDF). Proceedings of the Royal Society B 274 (1613): pp. 1117-1123. doi:. http://www.stanford.edu/~dmgordon/frederickson_gordon2007.pdf. Consultado el 30 de mayo de 2009. Resumen divulgativo – Suplemento Natura de «El Mundo» (11/3/2006).
- ↑ Beattie, Andrew J.; C. Turnbull, R. B. Knox y E. G. Williams (1984). «Ant Inhibition of Pollen Function: A Possible Reason Why Ant Pollination is Rare» (en inglés). American Journal of Botany 71 (3): pp. 421-426. doi:.
- ↑ Katayama N, Suzuki N (2004). «Role of extrafloral nectaries of Vicia faba in attraction of ants and herbivore exclusion by ants» (en inglés). Entomological Science 7 (2): pp. 119-124. doi:. http://onlinelibrary.wiley.com/doi/10.1111/j.1479-8298.2004.00057.x/pdf.
- ↑ Fischer RC, Wanek W, Richter A, Mayer V (2003). «Do ants feed plants? A 15N labelling study of nitrogen fluxes from ants to plants in the mutualism of Pheidole and Piper» (en inglés). Journal of Ecology 91: pp. 126-134. doi:.
- ↑ Hanzawa FM, Beattie AJ, Culver DC (1988). «Directed dispersal: demographic analysis of an ant-seed mutualism» (en inglés). American Naturalist 131 (1): pp. 1-13. doi:.
- ↑ Giladi I (2006). «Choosing benefits or partners: a review of the evidence for the evolution of myrmecochory» (en inglés). Oikos 112 (3): pp. 481-492. doi:.
- ↑ Lengyel, S. et ál (2010). «Convergent evolution of seed dispersal by ants, and phylogeny and biogeography in flowering plants: A global survey» (en inglés). Perspectives in Plant Ecology, Evolution and Systematics 12: pp. 43–55. doi:. http://ecology.science.unideb.hu/ConsEcolGroup/Pdf/Lengyel%20et%20al%202010%20(MyrmecochoryReview).pdf.
- ↑ Fischer RC, Ölzant SM, Wanek W, Mayer V (2005) (en inglés). The fate of Corydalis cava elaiosomes within an ant colony of Myrmica rubra: elaiosomes are preferentially fed to larvae. 52. pp. 55-62. doi:.
- ↑ Hughes L, Westoby M (1992). «Capitula on stick insect eggs and elaiosomes on seeds: convergent adaptations for burial by ants» (en inglés). Functional Ecology 6: pp. 642-648. doi:.
- ↑ Quinet Y, Tekule N y de Biseau JC (2005). «Behavioural Interactions Between Crematogaster brevispinosa rochai Forel (Hymenoptera: Formicidae) and Two Nasutitermes Species (Isoptera: Termitidae)» (en inglés). Journal of Insect Behavior 18 (1): pp. 1-17. doi:.
- ↑ Jeanne, RL (1972). «Social biology of the neotropical wasp Mischocyttarus drewseni» (en inglés). Bulletin of the Museum of Comparative Zoology (Harvard University) 144: pp. 63–150.
- ↑ a b Sivinski J, Marshall S, Petersson E (1999). «Kleptoparasitism and phoresy in the Diptera» (en inglés, PDF). Florida Entomologist 82 (2): pp. 179-197. doi:. http://www.fcla.edu/FlaEnt/fe82p179.pdf.
- ↑ Schaechter E (2000). «Some weird and wonderful fungi» (en inglés). Microbiology Today 27 (3): pp. 116-117.
- ↑ Sandra B. Andersen et ál. (2009). «The Life of a Dead Ant: The Expression of an Adaptive Extended Phenotype» (en inglés). The American Naturalist 174 (3): pp. 424-433. doi:.
- ↑ Wojcik DP (1989). «Behavioral interactions between ants and their parasites» (en inglés). The Florida Entomologist 72 (1): pp. 43-51. doi:.
- ↑ Poinar G Jr., Yanoviak SP (2008). «Myrmeconema neotropicum n. g., n. sp., a new tetradonematid nematode parasitising South American populations of Cephalotes atratus (Hymenoptera: Formicidae), with the discovery of an apparent parasite-induced host morph» (en inglés, PDF). Systematic Parasitology 69 (2): pp. 145-153. doi:. http://www.canopyants.com/2008_SystParasit.pdf.
- ↑ Caldwell JP (1996). «The evolution of myrmecophagy and its correlates in poison frogs (Family Dendrobatidae)» (en inglés). Journal of Zoology 240 (1): pp. 75-101. doi:.
- ↑ Willis, E. y Oniki Y. (1978). «Birds and Army Ants» (en inglés). Annual Review of Ecology and Systematics 9: pp. 243-263. doi:.
- ↑ Vellely AC (2001). «Foraging at army ant swarms by fifty bird species in the highlands of Costa Rica» (en inglés, PDF). Ornitologia Neotropical 12: pp. 271-275. http://www.ibiologia.unam.mx/pdf/links/neo/rev12/vol_12_3/orni_12_3_%20271-276.pdf.
- ↑ Wrege PH; Wikelski M, Mandel JT, Rassweiler T, Couzin ID (2005). «Antbirds parasitize foraging army ants» (en inglés). Ecology 86 (3): pp. 555-559. doi:.
- ↑ Swenson JE, Jansson A, Riig R, Sandegren R (1999). «Bears and ants: myrmecophagy by brown bears in central Scandinavia» (en inglés). Canadian Journal of Zoology 77 (4): pp. 551-561. doi:.
- ↑ «Silenciosas e incansables, nuevas especies de hormiga». Universidad Nacional del Noroeste de la Provincia de Buenos Aires (22/1/2009). Consultado el 18 de mayo de 2009.
- ↑ Juan Mousques (16/9/2007). «Hormigas cortadoras causan importantes pérdidas». Crónica Rural. Consultado el 18 de mayo de 2009.
- ↑ Gottrup F, Leaper D (2004). «Wound healing: Historical aspects» (en inglés, PDF). EWMA Journal 4 (2): pp. 21-26. http://ewma.org/fileadmin/user_upload/EWMA/pdf/journals/EWMA_Journal_Vol_4_No_2.pdf.
- ↑ Gudger EW (1925). «Stitching wounds with the mandibles of ants and beetles» (en inglés). Journal of the American Medical Association 84: pp. 1861-1864.
- ↑ Sapolsky, Robert M (2001) (en inglés). A Primate's Memoir: A Neuroscientist's Unconventional Life Among the Baboons. Simon and Schuster. p. 156. ISBN 0743202414.
- ↑ Haddad Jr. V, Cardoso JLC, Moraes RHP (2005). «Description of an injury in a human caused by a false tocandira (Dinoponera gigantea, Perty, 1833) with a revision on folkloric, pharmacological and clinical aspects of the giant ants of the genera Paraponera and Dinoponera (sub-family Ponerinae)» (en inglés, PDF (680 KB)). Revista do Instituto de Medicina Tropical de São Paulo 47 (4): pp. 235-238. doi:. http://www.scielo.br/pdf/rimtsp/v47n4/25664.pdf.
- ↑ McGain F, Winkel KD (2002). «Ant sting mortality in Australia» (en inglés). Toxicon 40 (8): pp. 1095-1100. doi:.
- ↑ Downes D, Laird SA (1999). «Innovative mechanisms for sharing benefits of biodiversity and related knowledge» (en inglés) (PDF). The Center for International Environmental Law. Consultado el 5 de mayo de 2009.
- ↑ Cheney RH, Scholtz E (1963). «Rooibos tea, a South African contribution to world beverages» (en inglés, PDF). Economic Botany 17 (3): pp. 186-194. doi:. ISSN 1874-9364.
- ↑ Chapman, RE y Bourke, Andrew FG (2001). «The influence of sociality on the conservation biology of social insects» (en inglés, PDF). Ecology Letters 4 (6): pp. 650-662. doi:. http://onlinelibrary.wiley.com/doi/10.1046/j.1461-0248.2001.00253.x/pdf.
- ↑ DeFoliart GR (1999). «Insects as food: Why the western attitude is important» (en inglés). Annual Review of Entomology 44: pp. 21-50. doi:.
- ↑ Bingham CT (1903) (en inglés). Fauna of British India. Hymenoptera Volume 3. p. 311.
- ↑ a b Bequaert J (1921). «Insects as food: How they have augmented the food supply of mankind in early and recent times» (en inglés). Natural History Journal 21: pp. 191-200.
- ↑ «Pest Notes: Ants» (en inglés). University of California Agriculture and Natural ResourcesHow to Manage Pests. Pests of Homes, Structures, People, and Pets (2007). Consultado el 7 de mayo de 2009.
- ↑ «Two step method for fire ant control» (en inglés). Oklahoma State University. Consultado el 7 de mayo de 2009.
- ↑ «100 of the World's Worst Invasive Alien Species» (en inglés y español). Invasive Species Specialist Group (ISSG). Consultado el 18 de julio de 2009.
- ↑ «Some general impacts of invasive ants» (en inglés) (PDF). Invasive Species Specialist Group (ISSG). Consultado el 18 de julio de 2009.
- ↑ De Bach, P. (1969). Control biológico de las plagas de insectos y malas hierbas. La Habana: Editorial Revolucionaria. p. 50.
- ↑ Che-Lung (1976). «Biological control of pest in China». Acta Entomológica Sinaica 19 (3): pp. 251-252.
- ↑ Rijo Camacho, E. «Control biológico del tetuán del boniato (Cylas formicarius elegantulus Sum.)». Aguascalientes (México): Fundación Produce. Consultado el 18 de julio de 2009.
- ↑ M. Tindo, P.S. Mbenoun, M. Kenne, J. Orivel, A.Dejean. «La petite fourmis de feu Wasmannia auropunctata (Roger) au Cameroun: distribution et structure de la colonie» (en francés). 24-27 Avril 2006 Colloque annuel UIEIS-SF. Union Internationale pour l'Etude des Insectes Sociaux -Section française- Université d'Avignon et des pays de Vaucluse, Avignon. http://www.avignon.inra.fr/content/download/3904/62572/file/UIEIS_Avignon-2006-R%E9sum%E9s.pdf.
- ↑ Holway, D.A.; Lach, L.; Suárez, A.V.; Tsutsui, N.D. y Case, T.J. (2002). «The causes and consequences of ant invasions» (en inglés, PDF). Annual Review of Ecology and Systematics 33: pp. 181-233. doi:. http://nature.berkeley.edu/tsutsuilab/Holwayetal_2002_ARES.pdf.
- ↑ Ness, J.H. y Bronstein, J.L. (2004). «The effects of invasive ants on prospective ant mutualists» (en inglés). Biological Invasions 6 (4): pp. 445-461. doi:.
- ↑ Lubbock, J (1881). «Observations on ants, bees, and wasps. IX. Color of flowers as an attraction to bees: Experiments and considerations thereon» (en inglés, PDF (6 MB)). Journal and Proceedings of the Linnean Society of London. Zoology 16: pp. 110–112. doi:. http://www.archive.org/download/cu31924003078163/cu31924003078163.pdf.
- ↑ Stadler B, Dixon, AFG (2008) (en inglés). Mutualism: Ants and their insect partners. Cambridge University Press. ISBN 978-0-521-86035-2.
- ↑ Kennedy CH (1951). «Myrmecological technique. IV. Collecting ants by rearing pupae» (en inglés). The Ohio Journal of Science 51 (1): pp. 17-20. https://kb.osu.edu/dspace/bitstream/handle/1811/3802/V51N01_017.pdf?sequence=1.
- ↑ Wojcik DP, Burges RJ, Blanton CM, Focks DA (2000). «An improved and quantified technique for marking individual fire ants (Hymenoptera: Formicidae)» (en inglés, PDF). The Florida Entomologist 83 (1): pp. 74-78. doi:. http://www.fcla.edu/FlaEnt/fe83p74.pdf.
- ↑ Dicke E, Byde A, Cliff D, Layzell P (2004) (en inglés). Proceedings of Biologically Inspired Approaches to Advanced Information Technology: First International Workshop, BioADIT 2004 LNCS 3141. pp. 364-379.
- ↑ Booker, L (verano 2005). «Learning from Nature: Applying Biomimetic Approaches to Military Tactics and Concepts» (en inglés, PDF). The Edge (MITRE Corporation) 9 (1): pp. 10-11. http://www.mitre.org/news/the_edge/spring_05/edge_spring_05.pdf.
- ↑ Arquilla, J y Ronfeldt, D (2000) (en inglés, PDF). Swarming and the Future of Conflict. Santa Mónica, California: RAND & National Defense Research Institute, United States Department of Defense. pp. 25-26. ISBN 0-8330-2885-5. http://www.rand.org/pubs/documented_briefings/2005/RAND_DB311.pdf.
- ↑ Corán 27:18–19.
- ↑ (en inglés) Sahih Bukhari. 4, libro 54, número 536. http://www.alquranic.com/articles/mail254.html.
- ↑ Deen, Mawil Y. Izzi (1990). «Islamic Environmental Ethics, Law, and Society». En Engel JR y Engel JG (en inglés, PDF). Ethics of Environment and Development. Bellhaven Press, Londres. http://www.mbcru.com/Texas%20Tech%20Mypage/Conservation%20Biology/Assignment%202/IzziDeenIslamicEcol.pdf.
- ↑ Balee WL (2000). «Antiquity of traditional ethnobiological knowledge in Amazonia: The Tupi-Guarani family and time» (en inglés). Ethnohistory 47 (2): pp. 399-422. doi:.
- ↑ Cesard N, Deturche J, Erikson P (2003). «Les Insectes dans les pratiques médicinales et rituelles d’Amazonie indigène». En E. Motte-Florac y J. M. C. Thomas (en francés, PDF). Les insectes dans la tradition orale. París: Peeters-Selaf. pp. 395-406. http://hal.archives-ouvertes.fr/docs/00/34/71/53/PDF/Insectes_Amazonie_indigene-CESARD-DETURCH_ERIKSON.pdf.
- ↑ Schmidt, R. J. (1985). «The super-nettles: a dermatologist's guide to ants in the plants» (en inglés). International Journal of Dermatology 24 (4): pp. 204-210. doi:. PMID 3891647. http://www.botanical-dermatology-database.info/BotDermReviews/Myrmecophytes.html.
- ↑ Twain, Mark (1880). «22 The Black Forest and Its Treasures» (en inglés). A Tramp Abroad. Nueva York: Oxford University Press. ISBN 0195101375. http://www.gutenberg.org/files/119/.
- ↑ Wilson, EO (25/1/2010). «Trailhead» (en inglés). The New Yorker. Consultado el 25 de junio de 2010.
- ↑ Guri, Assaf (8/9/1998). «Habitat media for ants and other invertebrates (US Patent 5803014)» (en inglés). United States Patent and Trademark Office. Consultado el 31 de diciembre de 2009.
- ↑ «1992 Excellence in Software Awards Winners» (en inglés). Software & Information Industry Association. Consultado el 11 de mayo de 2009.
- ↑ Rick Harbaugh (1998) (en inglés). Chinese Characters: A Genealogy and Dictionary. Yale University Press. ISBN 0966075005. http://zhongwen.com/cgi-bin/zipux2.cgi?b5=%C3%C6.
- ↑ Hearn L (1904) (en inglés, ASCII). Kwaidan: Stories and studies Of strange things. Tuttle publishing (reimpresión año 2005). p. 223. ISBN 0804836620. http://www.gutenberg.org/dirs/etext98/kwidn10.txt. Consultado el 30 de mayo de 2009.
- ↑ Sharkey, AJC (2006). «Robots, insects and swarm intelligence» (en inglés). Artificial Intelligence Review 26 (4): pp. 255-268. doi:.
Bibliografía
- Donal J. Borror, Charles A. Triplehorn, Norman F. Johnson (1989) (en inglés). Introduction to the Study of Insects (6ª edición). Saunders College Publishing. ISBN 0030253977.
- Bert Hölldobler, Edward O. Wilson (1990) (en inglés). The Ants. Harvard University Press. ISBN 0674040759. http://books.google.com/books?id=ljxV4h61vhUC&pg=PP1&dq=The+Ants&hl=es.
Lectura adicional
- Huber, Pierre (1867). Historia de las hormigas. Traducido por M. Fernández Llamazares. Imp. y Libr. de Gaspar y Roig, Eds. http://books.google.es/books?id=2RWv86vFQGwC&printsec=frontcover&source=gbs_v2_summary_r&cad=0#v=onepage&q=&f=false.
- Markle, Sandra (2007). Las hormigas legionarias. Animales Carroñeros. Ediciones Lerner. ISBN 0822577305. http://books.google.es/books?id=OGa3goXvEZUC&printsec=frontcover&source=gbs_v2_summary_r&cad=0#v=onepage&q=&f=false.
- Tanquary, Maurice Cole (2009). BiblioBazaar, LLC. ed (en inglés). Biological and Embryological Studies on Formicidae. ISBN 0559981015.
Enlaces externos
 Wikiespecies tiene un artículo sobre Formicidae. Wikispecies
Wikiespecies tiene un artículo sobre Formicidae. Wikispecies Wikimedia Commons alberga contenido multimedia sobre Formicidae. Commons
Wikimedia Commons alberga contenido multimedia sobre Formicidae. Commons- Hormigas.org, web sobre hormigas, con claves de identificación.
- divulgaUNED.es, artículo de divulgación científica sobre la racionalidad colectiva de las hormigas del género Temnothorax.
- Formicidae.org, página de desarrollo comunitario sobre las hormigas (en inglés).
- AntBase, base de datos taxonómica con abundante bibliografía (en inglés).
- Antmacroecology.org, mapas que muestran la distribución en el mundo de todos los géneros conocidos (en inglés).
Categorías:- Fósiles temporal
- Formicidae
- Plagas


























Wikimedia foundation. 2010.